INTRODUCTION
Working memory impairments are considered central in schizophrenia [1]. Because working memory impairments are one of the most enduring symptoms, they are also strong predictors of poor clinical outcome in individuals with schizophrenia [2]. Thus, individuals with schizophrenia often experience problems in their daily activities, such as difficulties remembering phone numbers or engaging in mental arithmetic. The causes behind working memory abnormality have been predominantly reported through the volume and thickness of gray matter (GM) [3]. Such studies, however, have only focused on the macrostructural differences and thus have not captured microstructural abnormalities. Diffusion tensor imaging (DTI) studies are useful to study microstructural abnormalities through white matter (WM), and WM-DTI studies have reported that the abnormalities of working memory are associated with WM [4-6] and depend on myelinated axonal fibers [7-10].
DTI studies have typically investigated WM microstructure by examining the water molecule’s diffusivity in the brain. Compared to WM-DTI studies, only a few studies have investigated GM microstructure in the frontal and/or temporal cortex [11-14], entorhinal cortex [15], and parahippocampal, insula, and anterior cingulate regions [16] of individuals with schizophrenia. These studies were conducted using diffusivity in GM, such as axial diffusivity (AD), radial diffusivity (RD), fractional anisotropy (FA), trace (TR) or mean diffusivity (MD). As a result, positive GM-DTI findings in schizophrenia have been reported: 1) increased MD was shown, bilaterally, in the superior temporal gyrus (STG) GM and the left STG WM. MD in left STG WM demonstrated the association with attentional impairments and auditory hallucinations in schizophrenia [12]; 2) increased TR in the bank of the superior temporal sulcus of left hemisphere and the inferior temporal, the middle temporal, the inferior parietal gyri of right hemisphere in schizophrenia [11]. TR in the middle temporal gyrus and the inferior temporal gyrus of right hemisphere had also negative correlation with social cognition in schizophrenia [11]; 3) increased AD and RD were shown in the temporal and prefrontal cortices. AD and RD were associate with positive symptom of schizophrenia [13]; 4) there is elevated diffusivity in the frontotemporal regions on schizophrenia [14]; 5) increased MD was showed in the insula, anterior cingulate gyrus, and parahippocampal gyrus of right hemisphere in schizophrenia [16].
The diffusion anisotropy in GM cortex has been observed in the animal [17-20], and the human [21-26]. Radial diffusion anisotropy has also been demonstrated in the animal [27,28], and the human [28-35]. The principal diffusion direction shows apparent structure within GM, running nearly perpendicular to the cortical surface [30]. And the principal diffusion component’s orientation tends to be more within a plane parallel to the cortical surface [36]. Although RD and AD have been focused on the evaluation of axon integrity in WM, evidence demonstrate that the direction of diffusion in GM occurs both radial (perpendicular) and axial (parallel) to the surface of the cortex [28,30]. Thus, RD and AD in GM may reflect pyramidal cell’s microstructural integrity [37]. Investigating this phenomenon, a detailed study suggests that parallel and radial structure are believed to exist, and actually be a microscopic marker that is able to distinguish brain areas [28].
The cause of lower cases of GM-DTI than WM-DTI is that GM studies have typically investigated volume and thickness reductions in schizophrenia. Interestingly, in individuals with schizophrenia, the decreased GM thickness and volume have been shown by reduced presence of membranes, axon terminals, dendrites, and dendritic spines [38-40]. These findings suggest that the intrinsic microstructure of GM, rather than its volume and thickness, is more important for understanding the neuropathological basis of GM abnormalities in schizophrenia.
Despite the advantages of the use of DTI to study GM microstructure, previous DTI studies have focused on WM, and the few GM-DTI studies have focused on specific GM regions or they involved small sample sizes. According to previous schizophrenia studies, there are more than 50 regional deficits, such as in frontal, temporal, parietal, and cingulate areas and in the deficits [41], and a larger group will give us better understanding the feature of these microscopic alternations and their associations with clinical profiles and symptoms [42]. Therefore, it is necessary to study whole GM regions with larger samples using sufficient data sources. The SchizConnect database (www.schizconnect.org) shows Significant possibilities to overcome the barriers related to the creation of large data sets to elevate statistical power [43]. Consequently, our study used the SchizConnect database to examine entire GM regions in a larger sample.
This study investigated the hypothesis that GM diffusivity in regions of interest (ROIs) would differ between individuals with schizophrenia and control subjects and that GM diffusivity and working memory would correlate significantly.
METHODS
Data sources
This study used the demographic data, brain images (T1 and DTI), and working memory performance test data of 229 cases included in the SchizConnect database used in the Center for Biomedical Research Excellence (COBRE) and Neuromorphometry by Computer Algorithm Chicago (NMorphCH) studies [44].
Study participants
Of the 229 cases, 42 were excluded for the following reasons: missing cognitive function test values (n=22), diagnoses of schizoaffective disorder (n=10), and poor brain imaging quality (n=10). The remaining 187 cases were divided into two groups: patients with schizophrenia (n=90) and control subjects (n=97). The age range of the participants was 20 to 66 years. This study was approved by the Institutional Review Board of Asan Medical Center (IRB File No. S2017-1996-0001).
Diffusivity
T1 and DTI neuroimages were used. In the COBRE database, the T1 scan parameters were as follows: repetition time (TR)=2,530 ms, echo time (TE)=1.64 ms, flip=0°, matrix=256×256, slices=192, and slice thickness=1 mm. The DTI scan parameters were as follows: TR=9,000 ms, TE=84 ms, flip=0°, matrix=256×256, slices=72, and slice thickness=2 mm. In the NMorphCH database, the T1 scan parameters were as follows: TR=2,400 ms, TE=3.16 ms, flip=8°, matrix=256×256, slices=176, and slice thickness=1 mm. The DTI scan parameters were as follows: TR=8,000 ms, TE=86 ms, flip=90°, matrix=896×896, slices=35, and slice thickness=2 mm.
Motion and eddy current-induced distortions were corrected with the affine registration of all gradient volumes with the first b=0 volume using the FMRIB Software Library (FLIRT: FSL, version 6.0.0; http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/) [45,46]. Our calculations of diffusivities were conducted using eigenvalues (λ1, λ2, and λ3). FA was automatically calculated in dtifit in FSL. The following additional diffusivity values were obtained for the whole brain of each subject using eigenvalues in fslmaths in FSL: 1) AD (λ1), 2) RD [(λ2 + λ3)/2], and 3) TR (λ1+λ2+λ3).
The T1 images were parcellated into discrete anatomical regions using the Desikan-Killiany atlas of FreeSurfer (version 6.0; http://surfer.nmr.mgh.harvard.edu) [47], and all parcellated ROIs of the GM (n=68) (Supplementary Table 1 in the online-only Data Supplement) were used in the subsequent analyses. The T1 images were registered to a b=0 baseline DTI image using flirt in FSL. The parcellated labels were then transformed into DTI using the same registration transformation. Next, we calculated the diffusivities (AD, FA, RD, and TR) for each transformed ROI of each subject. Z-scores were calculated using the means and standard deviations of each ROI of the control group in each project.
Symptoms
Patients’ psychiatric symptoms were evaluated by a psychiatrist using the Positive and Negative Syndrome Scale (PANSS) in COBRE and The Scale for the Assessment of Positive Symptoms (SAPS), the Scale for the Assessment of Negative Symptoms (SANS) in NMorphCH studies.
Working memory
We used the scaled scores from the Letter Number Sequencing (LNS) test. The LNS test assesses the subject’s ability to manipulate and rearrange verbal representations in working memory. The LNS test used in this study was included in the following different test batteries in the COBRE and NmorphCH projects: “Measurement and Treatment Research to Improve Cognition in Schizophrenia Consensus Cognitive Battery” [48,49] and “Wechsler Memory Scale Third Edition” [50], respectively. Thus, in this study, the scaled scores on the LNS test were converted into Z-scores for each project.
Statistical analysis
For the demographic data, t-tests were conducted to investigate differences in age, illness duration, and LNS scores between the groups. Chi-squared tests were conducted to examine differences in the sex ratios between the groups.
Next, statistical analyses controlling for sex and age were conducted. First, an analysis of covariance was conducted to test for significant differences in diffusivity (AD, FA, TR, and RD) in the 68 ROIs between the control and patient groups. Due to the multiple comparisons of diffusivity in the 68 ROIs, the significance threshold was adjusted using the false discovery rate (q<0.05). Second, Pearson partial correlation analyses were conducted on the LNS scores and ROI diffusivities showing significant differences between the groups. All statistical analyses were performed using R (R Foundation for Statistical Computing, Vienna, Austria) [51].
RESULTS
Demographic data
The demographic and clinical information is presented in Table 1. The final data set included 187 subjects (97 controls and 90 patients). The age of the groups did not differ significantly [mean±standard deviation (SD): controls, 36.74±11.56 years; schizophrenia, 36.94±11.44 years; df=184.23, t=-0.1201, p=0.90]. The percentage of men in the control group was 74.2%, higher than that in the schizophrenia group (72.2%), but this difference was not statistically significant (χ2=0.18, df=1, p=0.66). The mean duration of illness was 11.33±13.53 years in the individuals with schizophrenia in COBRE. The LNS scores differed significantly between the control and schizophrenia groups (controls: -0.02±1.01; schizophrenia: -0.99±1.26; df=170.65, t=5.82, p<0.001). PANSS total score was 59.54±19.89 (positive score: 15.24±5.25; negative score: 14.8±5.45; general score 29.5±9.19) in COBRE. SAPS total score was 42±34.39 and SANS total score was 53.31±26.27 in NMorphCH.
Comparisons of GM diffusivity
The diffusivity results are presented in Table 2. All p values were corrected by the false discovery rate. FA values did not differ significantly between the two groups (FDR-corrected p>0.01).
Regarding AD, the schizophrenia group exhibited significantly increased AD values compared with those of the control group in the following 3 ROIs: 1) left medial orbitofrontal cortex [mean±SD: control, 0.04±1.02; schizophrenia, 0.61±1.15; F(1, 183)=12.71, p<0.001], 2) left pars opercularis [control, 0.07±1; schizophrenia, 0.57±1.03; F(1, 183)=11.68, p<0.001], and 3) left STG [control, 0.08±0.98; schizophrenia, 0.54±1.1; F(1, 183)=9.66, p=0.002].
Regarding RD, the schizophrenia group exhibited significantly increased RD values compared with those of the control group in the following 14 ROIs: 1) left bank of the superior temporal sulcus [mean±SD: control, 0.08±0.99; schizophrenia, 0.5±0.98; F(1, 183)=8.3, p=0.004], 2) left inferior temporal gyrus [control, 0.08±0.99; schizophrenia, 0.5±0.98; F(1, 183)=9.61, p=0.002], 3) left lateral orbitofrontal cortex [control, 0.03±1.04; schizophrenia, 0.63±1.06; F(1, 183)=15.91, p<0.001], 4) left medial orbitofrontal cortex [control, 0.01±1.03; schizophrenia, 0.61±1.2; F(1, 183)=13.36, p<0.001], 5) left middle temporal gyrus [control, 0.01±1.04; schizophrenia, 0.59±1.02; F(1, 183)=16.59, p<0.001], 6) left pars opercularis [control, 0.04±1.02; schizophrenia, 0.71±1.1; F(1, 183)=20.1, p<0.001], 7) left rostral anterior cingulate cortex [control, 0.03±1.04; schizophrenia, 0.48±1.09; F(1, 183)=8.57, p=0.003], 8) rostral middle frontal gyrus [control, 0.03±1.02; schizophrenia, 0.46±0.86; F(1, 183)=10.39, p=0.001], 9) left STG [control, 0.04±1.03; schizophrenia, 0.59±0.98; F(1, 183)=15.38, p<0.001], 10) left temporal pole [control, 0.01±1.01; schizophrenia, 0.49±1.23; F(1, 183)=8.24, p=0.004], 11) right isthmus cingulate gyrus [control, 0.06±1.01; schizophrenia, 0.47±1.1; F(1, 183)=6.9, p=0.009], 12) right lateral orbitofrontal cortex [control, 0.03±1.02; schizophrenia, 0.47±1.19; F(1, 183)=7.2, p=0.008], 13) right pars opercularis [control, 0.05±1.03; schizophrenia, 0.49±1.29; F(1, 183)=7.18, p=0.008], and 14) right frontal pole [control, 0.01±1.03; schizophrenia, 0.47±1.2; F(1, 183)=8.84, p=0.003].
Regarding TR, the schizophrenia group exhibited significantly increased TR values compared with those of the control group in the following 9 ROIs: 1) left lateral orbitofrontal cortex [mean±SD: control, 0.03±1.04; schizophrenia, 0.56±1.05; F(1, 183)=12.03, p<0.001], 2) left medial orbitofrontal cortex [control, 0.02±1.02; schizophrenia, 0.63±1.2; F(1, 183)=13.95, p<0.001], 3) left middle temporal gyrus [control, 0.02±1.03; schizophrenia, 0.58±1.06; F(1, 183)=14.82, p<0.001], 4) left pars opercularis [control, 0.05±1.01; schizophrenia, 0.68±1.1; F(1, 183)=17.45, p<0.001], 5) left rostral anterior cingulate cortex [control, 0.04±1.04; schizophrenia, 0.48±1.09; F(1, 183)=8.36, p=0.004], 6) left rostral middle frontal gyrus [control, 0.04±1.01; schizophrenia, 0.45±0.89; F(1, 183)=9.11, p=0.002], 7) left STG [control, 0.06±1.01; schizophrenia, 0.59±1.02; F(1, 183)=14.27, p<0.001], 8) left temporal pole [control, 0.01±1.01; schizophrenia, 0.47±1.22; F(1, 183)=7.8, p=0.005], and 9) right frontal pole [control, 0.01±0.5; schizophrenia, 1.03±1.23; F(1, 183)=9.15, p=0.002].
Relationship between GM diffusivity and working memory
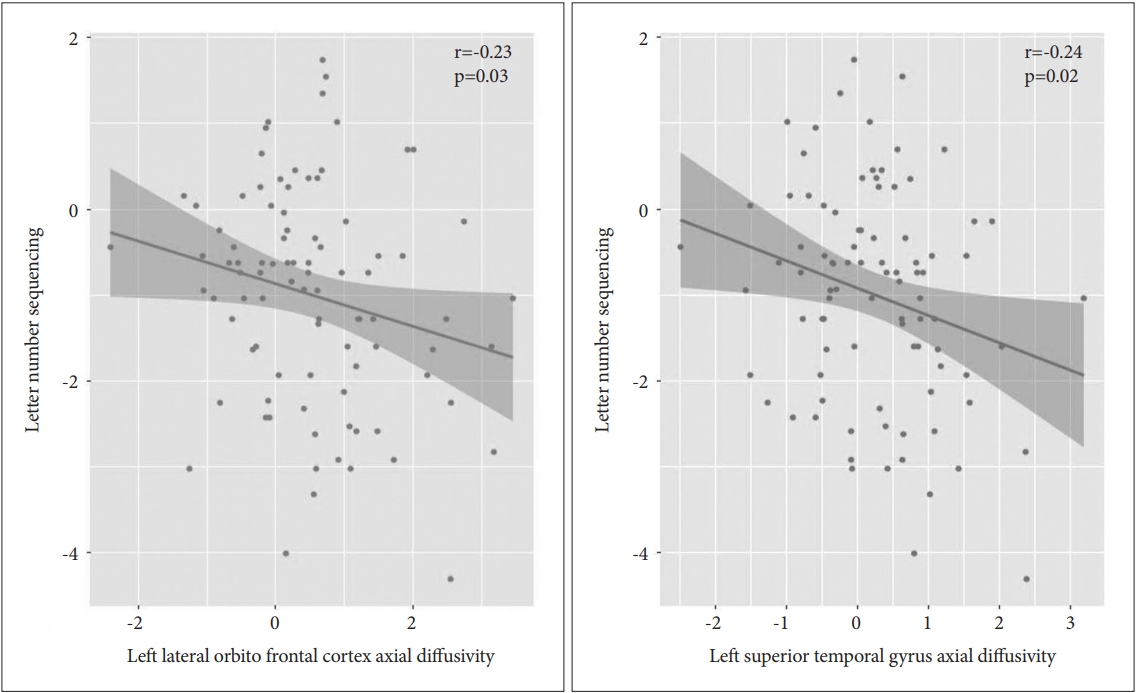
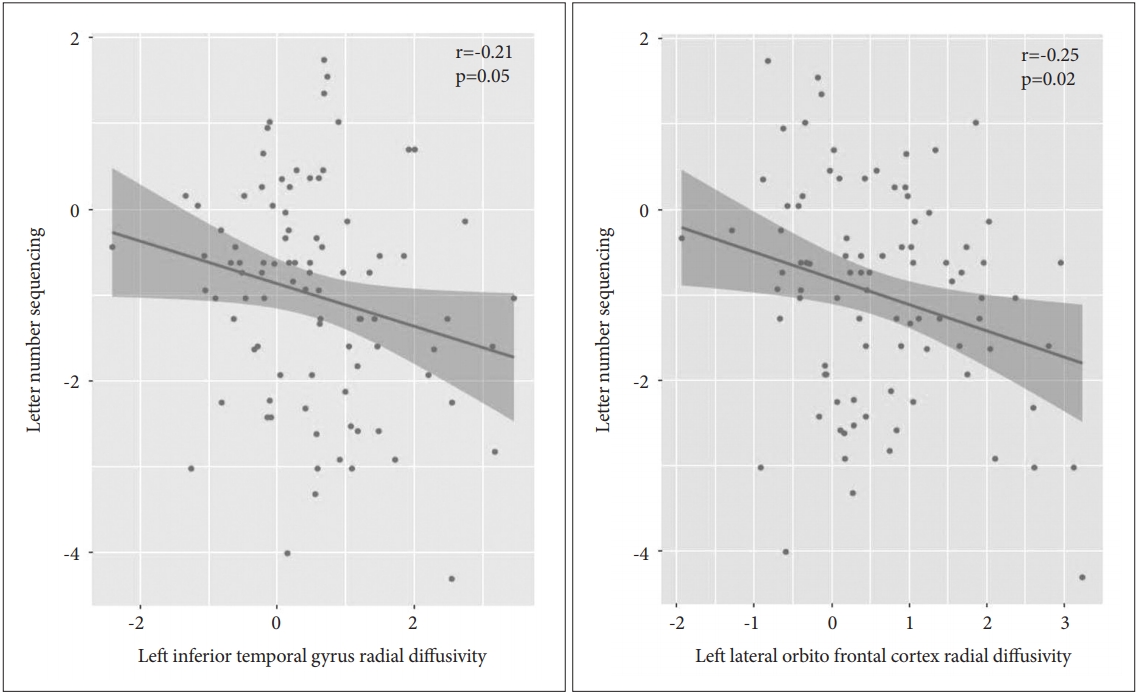
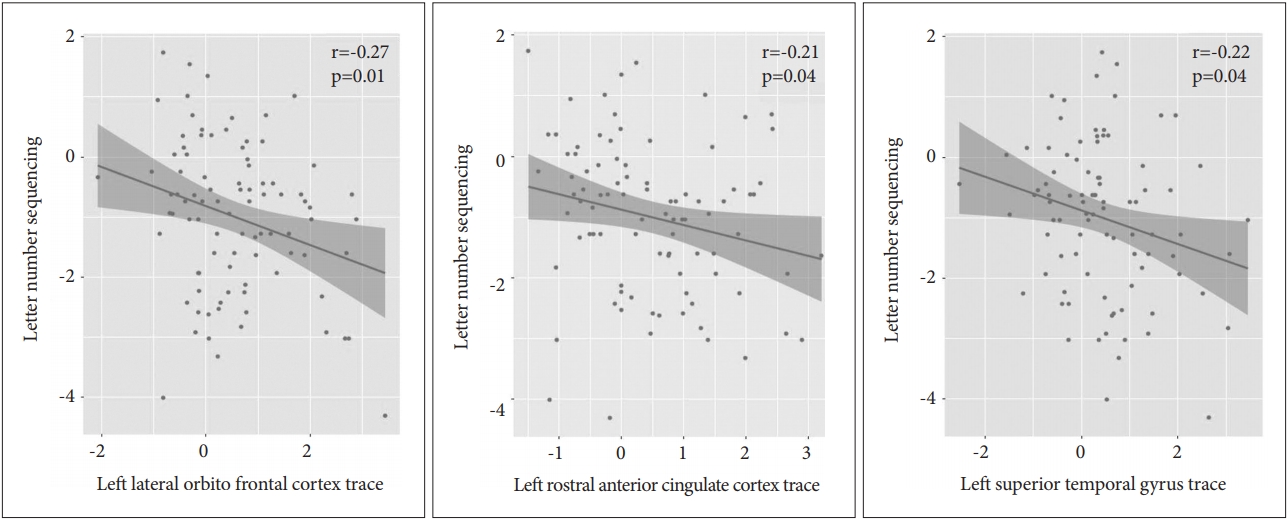
The correlation results are displayed in Table 3. In the patient group, we evaluated the Pearson partial correlations between the LNS scaled scores and the diffusivities (AD, RD, and TR) of the ROIs with diffusivity values that differed significantly from those of the control group. The AD values were significantly and negatively correlated with the LNS scores in the following ROIs (correlation coefficients, uncorrected p value) (Figure 1): 1) Left lateral orbitofrontal cortex (r=-0.23, p=0.03) and 2) Left STG (r=-0.24, p=0.02). The RD values significantly and negatively correlated with the LNS scores in the following ROIs (Figure 2): 1) Left inferior temporal gyrus (r=-0.21, p=0.05) and 2) Left lateral orbitofrontal cortex (r=-0.25, p=0.02). The TR values were significantly and negatively correlated with the LNS scores in the following ROIs (Figure 3): 1) left lateral orbitofrontal cortex (r=-0.27, p=0.01), 2) left rostral anterior cingulate cortex (r=-0.21, p=0.04), and 3) left STG (r=-0.22, p=0.04).
DISCUSSION
The purpose of this study was to compare the GM microstructures between the schizophrenia and control groups and investigate the relationship between GM diffusivity and working memory. Compared with the control group, the schizophrenia group showed significantly higher AD, RD, and TR values in specific regions in frontal, temporal, and anterior cingulate regions. Working memory and diffusivity (AD, RD, and TR) were negatively correlated in the lateral orbitofrontal, superior temporal, inferior temporal, and rostral anterior cingulate areas of the left hemisphere in the individuals with schizophrenia.
Previous GM-DTI studies have shown that, compared with controls, individuals with schizophrenia have increased AD and RD values in the prefrontal cortex [13], increased AD, RD, and TR values in the left bank of the superior temporal sulcus [11]; and increased MD, similar to TR, in the STG [12]. Our results were partly consistent with the previous results. Increased RD and TR values indicate microstructural abnormalities that suggest alterations in intrinsic connections [52], a loss of dendritic spines [38,53,54], and/or increased volume of extracellular water due to inflammation [55-57]. No significant changes were found in the FA but were found in the TR because FA measures directionality and TR measures the proportion of water movement. Thus, TR, and FA can be independent of each other [58]. Therefore, the findings of increased TR and no change in FA might be associated with increased water content as in cases of edema [59] or a subtle loss of neurological processes [39,52,60]. Interestingly, the AD values were increased in the schizophrenia group, which differed from the findings of previous DTI studies on schizophrenia. However, several DTI-schizophrenia studies have reported increased AD values and suggested that decreases or delays in the normal pruning process or neurodegenerative processes may lead to axonal loss [61] and increased water content outside of the axonal space [62]. These findings suggest that increased AD is associated with neurological issues.
The previous studies investigating the association between the brain structure and working memory have deemed that the orbitofrontal cortex is necessary for the working memory maintenance, manipulation, and monitoring processes, and high levels of cognitive load [63,64]. LNS is an appropriate test of such conditions. LNS requires the ability to monitor a series of complex stimuli, maintain recently processed the items, and manipulates items in a series to identify correct sorting. The results of our study support those of a previous study that reported an association between the orbitofrontal cortex and performance on working memory tasks requiring high levels of cognitive load, such as the LNS. Thus, considering our results were indicative of increased diffusivity (RD and TR) in the orbitofrontal cortex, microstructural abnormalities in the orbitofrontal cortex might serve as an indicator of the processing of the coordination of the working memory maintenance, manipulation, and monitoring processes.
Abnormalities in the anterior cingulate cortex, which is a key component of working memory, are strongly associated with schizophrenia [65-67]. Several studies have reported that abnormal synaptic connectivity and decreased neuronal density in the anterior cingulate cortex may contribute to working memory impairments in schizophrenia [68-71]. In addition, the rostral and caudal anterior cingulate cortices share inputs with a network involving associative, limbic, sensorimotor, dorsolateral prefrontal area, which is a structure that is independently linked to working memory [72]. We found negative correlations between working memory and diffusivity, indicating microstructural abnormalities in the anterior cingulate cortex. Based on previous findings, our results suggest that working memory problems are related to difficulty in the sharing of information with each area that is connected to the anterior cingulate cortex due to microstructural abnormalities, such as synaptic dysconnectivity and decreased neuronal density in the anterior cingulate cortex.
Temporal areas are consistently activated during working memory performance [73]. In addition, the left inferior temporal and left superior temporal gyri are associated with short-term memory [74,75]. Previous studies with patients with shortterm memory impairments have reported that decreased verbal working memory is due to damage to verbal shortterm [76-78]. Considering these previous results and our results of the negative correlation between working memory and diffusivity in temporal areas, we suggest that microstructural deficits in temporal areas result in poor working performance and are therefore associated with short-term memory deficits for working memory load.
The strength of this study is that we studied the whole GM region with more than 150 participants using a large database. According to previous schizophrenia studies, there are more than 50 regional deficits, such as frontal, temporal, parietal, cingulate area and cuneus deficits [41], and a larger group size will give us the specificity of microscopic alterations and the implication that the alterations are associated with symptoms, or clinical profiles [42]. Previous GM-DTI studies were conducted within a specific GM region or small sample size. Such studies may ignore GM regions that are important from a clinical perspective or require more explanatory power. Therefore, a GM-DTI study of the whole GM region with a larger sample size is meaningful to increase the explanatory power of GM microstructural abnormalities.
Our study had the following limitations. First, we did not measure other cognitive functions because only the scale scores for the LNS test were available in the datasets of the two different research projects. Future studies should investigate the associations of other cognitive functions with GM diffusivity. Second, this study used data from two different projects. Thus, our study was conducted on data that were obtained from different neuroimaging systems and different cognitive batteries. Therefore, standard scores were used within the framework of our study. Future studies need to be conducted using the same protocol and similar sample sizes. Third, we did not consider illness duration or the duration of medications in this study because the two research projects did not contain the same clinical data. Future studies need to be conducted for comparisons to the results of this study, especially in consideration of these variables.
Our results indicated GM microstructural abnormalities in the frontal, temporal, and anterior cingulate regions of individuals with schizophrenia. Furthermore, these regional GM microstructural abnormalities suggest a neuropathological basis for the working memory deficits clinically observed in individuals with schizophrenia.