INTRODUCTION
Affective temperaments refer to characteristic patterns in the quality and intensity of baseline mood levels and emotional and behavioral responses and regulations to internal and external stimuli, which tend to appear early in life and be temporally stable [1-3]. Akiskal et al. [1] proposed five affective temperamental traits: depressive, cyclothymic, hyperthymic, irritable, and anxious temperaments and developed a valid assessment scale for affective temperaments, viz. the Temperament Evaluation of the Memphis, Pisa, Paris, and San Diego Autoquestionnaire (TEMPS-A). These affective temperaments are not only considered to be subclinical manifestation of affective disorders [1,4,5], but are also known to play a crucial role in the clinical prognosis of psychiatric illnesses [6-9]. A recent study of affective temperament and suicide showed that an anxious temperament is a strong risk factor for suicidal attempts, regardless of the presence of mental disorders, while depressive and irritable temperaments are risk factors for suicidal attempts in patients with mental disorders [6]. Therefore, a better understanding of affective temperaments is crucial to the understanding of the development of and treatments for mental disorders.
Affective temperaments are known to be moderately heritable, with heritability ranging from 21% to 52% for the five affective temperaments in 101 bipolar families [10]. Additionally, several candidate gene association studies have been conducted to identify genes for affective temperaments. Previous genetic association approaches of affective temperaments usually targeted monoaminergic systems, including serotoninrelated genes [11-14], and dopaminergic genes [13]. A genome-wide association study has revealed several variants that are significantly associated with a specific affective temperament in bipolar patients [15]. Since affective temperaments may be quantitative endophenotypes for affective disorders, representing genetic heterogeneity, the interest in genetic studies of affective temperaments has increased.
Besides the monoaminergic genes, the ubiquitin-proteasome system (UPS), a major protein degradation pathway, which is responsible for dynamic cellular processes, such as the regulation of cell cycle and the response to genotoxic and proteotoxic stress [16], may be a potential candidate system contributing to affective temperaments as well as affective disorders. The UPS is a critical contributor to the construction of brain networks via extensive regulation of synaptic plasticity [17,18]. In recent years, synaptic plasticity has emerged as a component of the pathophysiology of depression [19,20]. Interestingly, some studies have revealed direct associations between the UPS and depressive disorders [21-23]. In an animal study using quantitative trait locus mapping, the ubiquitin-specific peptidase 46 gene (USP46) was proposed to be responsible for depressive-like behavior in mice in depression-inducing tests, such as tail suspension and forced swimming tests [23]. Furthermore, USP46 knock-out mice also showed differences in depression-inducing test [22]. Moreover, a human study showed significant differences in USP46 single-nucleotide polymorphisms (SNPs) between patients with depressive disorder and normal controls [21]. Taken together, these findings may indicate that USP46 may be a candidate gene involved in the development of affective temperaments, particularly in depressive temperaments, as a subclinical manifestation of affective disorders.
The objective of the present study was to investigate the association of USP46 polymorphisms with affective temperamental traits in healthy subjects.
METHODS
Subjects
A total of 557 healthy Korean volunteers (334 males, 223 females) were recruited using advertisements. Most of the subjects were college students. All participants were interviewed by psychiatrists, and subjects with any current or previous Axis I psychiatric disorders according to the Diagnostic and Statistical Manual of Mental Disorders, 4th Edition (DSM-IV), a family history of psychiatric disorders, or neurologic disorders were excluded. Twelve subjects (2.16% of the total subjects) were excluded based on the exclusion criteria. A total of 545 subjects (328 male, 217 female) were included in the final analysis. All subjects provided informed consent before the study. The study protocol was approved by the Institutional Review Board of Severance Hospital (Severance IRB 4-2010-0577).
SNP selection
To date, only one study has reported positive association between USP46 gene and psychiatric phenotypes. Fukuo et al. [21] found significant association between haplotypes consisting of three (rs2244291-rs10034164-rs346005) or four (rs12646800-rs2244291-rs10034164-rs346005) SNPs of seven tagging SNPs (r2âĽ0.8) capturing 30 SNPs (minor allele frequency; MAFâĽ5) covering the USP46 gene region [21]. Therefore, we selected SNPs of rs12646800, rs2244291, rs10034164, and rs346005 in this study. However, we further excluded rs12646800 and rs10034164, as MAFs were less than 0.1 (0.056 for rs12646800, 0.087 for rs10034164). Especially, there were no minor homozygotes for rs10034164 in the HapMap database (release #27, population: Japanese in Tokyo). Finally, we included two SNPs (rs2244291 and rs346005) in this study. SNP rs2244291 is located in intron, and SNP rs346005 is located 2 kb upstream of USP46 start site (USP46; 52590960-52659335, rs346005; 52588572).
Genotyping
The DNA of the subjects was isolated from saliva collected in OG-500 OrageneÂŽ self-collection kit (tube format âOG-500â), using standard techniques. Genotyping of USP46 SNPs was performed using a single-base primer extension assay (ABI PRISMÂŽ SNaPShotTM Multiplex kit; ABI, Foster City, CA, USA) according to the manufacturerâs instructions. Genotyping was performed by DNA Link, Inc. (Seoul, South Korea), in a manner blinded to any other study data.
Assessments
Affective temperament was assessed with the Korean version of the Memphis, Pisa, Paris, and San Diego Autoquestionnaire (TEMPS-A, a self-reported temperament scale for evaluating temperamental dimensions, with five subscales, including depressive, cyclothymic, hyperthymic, irritable, and anxious temperaments [1]. This scale consists of 110 âyes-or-noâ type questions in total, of which 109 are applicable to male subjects. All participants completed the questionnaire.
Statistical analyses
Previous combined analysis showed the differences in TEMPSA subscales scores between males and females [24]. Therefore, all the analyses in this study were conducted separately by gender. Multiple analyses of covariance (MANCOVA) were performed using the total scores for each TEMPS-A affective temperament dimension as dependent variables, and USP46 SNP genotypes as fixed factors. Age was controlled as a covariate to control possible effects. The two SNPs were analyzed separately for each gender. Further analyses were conducted for significant associations, with Fisherâs least significant difference post-hoc tests to evaluate differences among genotypes. Data analysis was performed using SPSS 23.0 for Windows (IBM Corp., Armonk, NY, USA). The statistical significance was set at p<0.05 for all tests.
RESULTS
Baseline characteristics
Baseline demographic data and characteristics, including TEMPS-A scores, are summarized in Table 1. The mean age of the participants was 22.55 years (standard deviation; SD=2.64 years) for males and 21.57 years (SD=2.47 years) for females. The mean education level of the participants were 14.32 years (SD=1.46 years) for males and 14.39 years (SD=1.58 years) for females. There were significant differences in age (p<0.001) and depressive (p=0.003), cyclothymic (p=0.003), and anxious (p<0.001) dimension scores between male subjects and female subjects.
Association analyses between USP46 SNPs and five affective temperamental dimensions
The genotype distribution of the two USP46 SNPs, rs346005 and rs2244291 were in accordance with the Hardy-Weinberg equilibrium (Ď2=0.068, p=0.794 for rs346005, Ď2=0.161, p=0.6835 for rs2244291).
MANCOVA results are presented in Table 2 through Table 3. The MANCOVA showed a significant association between TEMPS-A and rs346005 genotypes in male subjects (Wilks Ν=0.929, F(10,640)=2.384, p=0.009). Among temperament dimensions, there was a significant difference in depressive temperament among the genotypes of rs346005 in male subjects (F=7.79, p<0.001, Ρp2=0.046). The association was not observed in female subjects (F=0.75, p=0.472, Ρp2=0.007) (Table 2). In contrast, for rs2244291, no associations were found between the genotypes and the five affective temperaments, in males or females (Table 3).
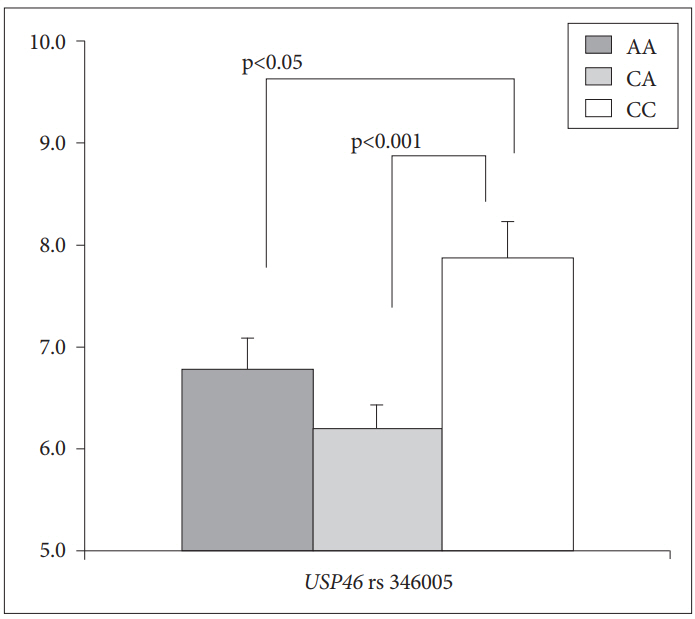
Post-hoc comparisons showed that subjects who had the CC genotype (mean=7.89, SD=0.45) at rs346005 showed a higher depressive temperament score than that of subjects with the AA genotype (mean=6.80, SD=0.30, p<0.05) and the CA genotype (mean=6.18, SD=0.21, p<0.0001) in male subjects. There is no significant difference in the depressive temperament score between subjects with the AA genotype and subjects with the CA genotype (p=0.103) (Figure 1).
DISCUSSION
We investigated the associations of the USP46 polymorphisms with the affective temperamental traits in healthy college students. The major finding was that the CC genotype of USP46 rs346005 was related to a higher depressive temperament score in male subjects. These findings suggest that affective temperamental traits are influenced by USP46.
Genetic associations between the ubiquitin proteasome system and affective temperament have not yet been reported. Although there is little direct evidence, our finding of the involvement of USP46 in depressive temperament is supported by indirect evidence. Theoretically, deubiquitinating enzymes, such as USP46, could play a role in regulating the stability of cellular protein and synaptic trafficking [25]. Animal studies have shown that USP46-mutated mice exhibit a significantly shorter immobility time than wild type mice in a tail suspension test (TST), which indicates a change in depressive state [23]. Furthermore, a genetic association study of major depression showed significant involvement of a USP46 haplotype in a Japanese population with major depressive disorder [21]. In the study, rs346005 was the only SNP that showed signification association with major depressive disorder. Considering that a depressive temperament is a subclinical form of major depressive disorder, it is possible that USP46 may be involved in the pathophysiology of depression.
The biological mechanism underlying the association between USP46 and depressive temperament remains unclear. However, some possible explanations of the association can be proposed. First, USP46 may act as a mood regulator by modulating neurotransmitter systems, such as serotonin and GABA. The antidepressant imipramine, a tricyclic antidepressant that inhibits reuptake of monoamines, such as serotonin and norepinephrine, reduced immobility in the TST of USP46 mutated mice [23], which supports the monoamine hypothesis [26]. In contrast, another study using USP46-knock-out mice reported that depressive behaviors in these mice were changed by administration of nitrazepam, GABAA receptor agonist [22], supporting the GABA hypothesis of major depressive disorder [27]. Second, the ubiquitin system is known to regulate the circadian system by controlling circadian clock protein [28]. Associations between the circadian system and affective temperament have previously been reported [29,30]. In a previous study, a depressive temperamental trait appeared to be associated with evening-type circadian preferences, along with cyclothymic, irritable, and anxious temperaments [30]. Therefore, it can be assumed that USP46 influences affective temperamental traits by modulating the circadian system. Third, USP46 may affect the affective temperament by modulating synaptic plasticity. USP46 has been reported to play a role in regulating alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propinic acid receptor (AMPAR), one of the key receptors involved in longterm potentiation and depression [25]. USP46, which is known to be enriched at the neural synapse, may modulate brain function by deubiquitinating AMPAR. Because evidence supporting these speculations is lacking, further studies elucidating the pathways that may mediate the association between UPS and depressive temperament should be performed.
Unlike in male subjects, no main effects of USP46 polymorphisms were observed in female subjects. This finding could be explained in several ways. In general, gene expression and splicing is known to differ between males and females [31]. From other perspectives, this gender difference might be related to an effect of sex hormones. Several studies have reported that the sex hormone estrogen modulates the ubiquitin systems [32,33], and plays an important role in synaptic plasticity [34]. In this study, there was no difference in genotype distribution of rs346005 between males and females. This suggested that USP46 rs346005 is not the major cause of difference in depressive temperament between males and females. In addition, considering the fact that complex traits, such as affective temperaments, are medicated by many genes (not one gene) and their interactions, the higher score of depressive temperament in females seems to be affected by other conditions, such as sexual hormone and/or X-chromosome. We could not find other studies showing the sexual dimorphisms of USP46 rs346005, since there is only a few existing studies on USP46. However, we found several other evidence, although not USP46, suggesting genetic sexual dimorphisms on temperamental traits. Stankova et al. [35] reported that the sexual dimorphic effects of oxytocin receptor gene (OXTR) showed gender difference on harm avoidance; one of the aspects of temperamental traits. In our previous study of dopamine receptor D4 (DRD4) gene on affective temperament, the association between DRD4 polymorphism and affective temperament (cyclothymic and irritable) was found only in male subjects [13]. In addition, since this study involved a small number of female subjects (n=217), the small sample size may not have been sufficient to obtain significant results.
There are several limitations in this study. First, most of our data were collected from college students, and it may not be generalizable. Although our sample can be considered a homogeneous group, with similar age, educational levels, and ethnicity, the results of this study should be considered cautiously. The results need to be replicated in other populations of different ages and ethnicities. Second, the present study was performed in a relatively small number of subjects. A further study in a larger sample size is needed to confirm the results. Third, we assessed only two SNPs. There are many more SNPs in USP46, and thus a more precise result could be obtained if additional SNPs were to be assessed. Fourth, affective temperamental scores could have been biased by social desirability, since TEMPS-A is a self-reporting scale. Fifth, we did not analyze gene-gene interaction and gene-environment interaction. Despite these limitations, this is the first study that reveals a possible genetic association between the UPS and affective temperaments.
In summary, the present study showed an association between a SNP in USP46 and depressive temperament. Our findings suggest that that some affective temperaments may be genetically predisposed to developing into affective disorders via vulnerable genes related to the UPS. In particular, USP46 may play a role in the pathophysiology of major depressive disorders by modulating a depressive temperament through dynamic proteolysis processes involving the UPS. Further research is required to elucidate influence of USP46 polymorphisms of affective temperaments in patients with mood disorders.