INTRODUCTION
The association between circadian rhythm and mood regulation has been established and disturbances in circadian clocks are suggested to contribute to the development of mood disorders [1-3]. The polymorphisms in the circadian genes, such as CLOCK, NPAS2, ARNTL1, NR1D1, PER3, RORB, and CSNK1, have been reported to be associated with mood disorders [4-6]. The drugs used to treat mood disorders affect the circadian rhythms and circadian genes [7-11]. The therapeutic modalities affecting the circadian clock have been applied to the treatment of mood disorders, such as light exposure and sleep-wake schedules shifting [11-14].
In mammals, circadian rhythms driven by the central pacemaker located in the suprachiasmatic nucleus (SCN) determine endogenous periodicity throughout the body [15]. Although the important role of circadian gene system in mood regulation has been suggested, there is little evidence to indicate the role of SCN in the pathogenesis of mood disorders. The extrahypothalamic brain regions including frontal cortex, cerebellum, and hippocampus have been reported to contain a circadian oscillator and to express all core clock genes [16-18]. The association between mood disorders and disrupted clock gene expression has been implicated in extra-SCN brain areas showing rhythmicity, such as medial prefrontal cortex, hippocampus, and amygdala [19-21].
The circadian clock is maintained and reset through a transcriptional feedback loop of core circadian genes [22]. Bmal1 and Clock dimerize and activate the transcription of Per and Cry; in turn, Per and Cry suppress their own expression through direct interaction with Bmal1/Clock dimers. In another feedback loop, Ror and Rev-erb, members of a subfamily of orphan nuclear receptors, directly regulate the expression of Bmal1 and Clock through the RORE motif, and Bmal1/Clock dimers reversely activate their expression [15,22]. Post-translational modifications also play an important role in regulation of the circadian clock system [23].
The electroconvulsive therapy (ECT) has been widely used to treat psychiatric disorders including mood disorders, which could correct the circadian and homeostatic abnormalities present in mood disorders [24]. It has been reported that single electroconvulsive seizure (ECS) treatment shows the enhanced Per1 mRNA expression [25] but repeated ECS treatment reduced it in the hippocampus [26]. ECS affects the expression of immediate early genes (IEGs) such as c-fos and early growth response protein1 (Egr1) in the hippocampus and frontal cortex [27-30], which induce changes in circadian gene expression [31,32], and modulates the activity of signaling transduction pathways, such as the protein kinase C (PKC), extracellular signal-regulated kinase (ERK) and glycogen synthase kinase-3β (GSK-3β) [33-35], involved in post-translation modifications of circadian proteins [23].
ECT is one of the major therapeutic modalities for the treatment of mood disorders, and the important role of circadian gene system in various brain regions in the pathogenesis and treatment of mood disorders has been increasingly understood. However, the investigations of the effects of ECT on circadian genes in the brain including extra-SCN regions are limited. Frontal cortex is one of the main regions involved in the regulation of mood-related behaviors [36,37], and the clock genes are expressed in all six layers of frontal cortex [17]. In this study, we examined the gene expression of core circadian clock components in the rat frontal cortex after single and repeated ECS treatments.
METHODS
Animals
Animals were treated in accordance with the National Institute of Health’s Guide for the Care and Use of Laboratory Animals, and formal approval to conduct this experiment was obtained from the Animal Subjects Review Board of Dongguk University Hospital. Male Sprague-Dawley rats (150-200 g) were housed for 1 week before the experiments and maintained under a strict 12-hr light/12-hr dark cycle (lights on 08:00hr), with food and water available ad libitum.
Electroconvulsive seizure treatment
ECS was applied in the rats using a previously validated technique [28,30]. ECS was administered via ear-clip electrodes (Ugo Basile ECT Unit-57800-001, Ugo Basile, Italy. Frequency 100 pulses/s, pulse width 0.5 ms, shock duration 0.5 s, current 55 mA). ECS-induced seizure was validated by the observation of general convulsion consisting of tonic and clonic phase and the time measurement of duration of convulsion (more than 30 s). Sham-treated control animals were handled in the same fashion as the ECS treatment group, but no electric current was delivered. Rats in the sham group underwent sham treatment for 10 days. Rats in the single-ECS treatment group (E1X) underwent sham treatment for 9 days and were then administered a single ECS treatment on day 10, whereas rats in the repeated-ECS treatment group (E10X) were treated with ECS for 10 days. All treatments were administered once a day at the same time (12:00-13:00). Male Sprague-Dawley rats with 6 weeks of age were started to undergo ECS or sham treatment as described above, and, at 8 weeks of age after ECS or sham treatments for 10 days, rats were decapitated at 24 hr after the last ECS treatment to determine the expression level at 24 hr. To determine the daily oscillation of circadian genes, rats were decapitated at 4, 8, 12, 16, 20, and 24 hr after the last treatment for the ZT8, ZT12, ZT16, ZT20, ZT0, and ZT4 samples, respectively, where ZT indicates the zeitgeber time. Decapitations in the dark cycle (ZT12-ZT0) were performed under a dim red light to prevent rats from additional light stimuli. The frontal cortex was used for further analyses.
Immunoblot analysis
Immunoblot analyses were performed as previously described [28]. Antibodies against Bmal1 (kindly gifted by Dr. Kyung Jin Kim, Seoul National University), Rev-Erbα (Cell Signaling Technology, Beverly, MA, USA), Rorα, Clock, Per1, Per2 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and β-actin (Sigma-Aldrich, St. Louis, MO, USA) were used at dilutions of 1:1000-1:5000. Blots were incubated with primary antibody overnight at 4°C. The membranes were subsequently incubated with HRP-conjugated secondary antibody (Zymed Laboratories, San Francisco, CA, USA), and the signals were detected using an enhanced chemiluminescence system (Pierce, Rockford, IL, USA). Immunoblot signals were quantified with the TINA program, version 2.10 g (Raytest, Straubenhardt, Germany). At least four independent experiments were performed.
Quantitative real-time PCR
Whole RNA was extracted from the frontal cortex using TRI reagent (Molecular Research Center, Cincinnati, OH, USA) and subsequently reverse-transcribed to cDNA using the Superscript II RT system (Invitrogen, Carlsbad, CA, USA). Quantitative real-time PCR was performed using an ABI PRISM 7500 instrument (Applied Biosystems, Foster City, CA, USA) with SYBR Green Real-time PCR master mix (Toyobo, Osaka, Japan) as previously described [28]. Relative amounts of all mRNAs were calculated by the comparative Ct method (Applied Biosystems) using the 2-ΔCt equation. ΔCt was the difference between the Ct values derived from the target gene and the β-actin control, and ΔΔCt was calculated by the formula ΔCt=ΔCt−ΔCt of the control group (sham or sham of CT8). The primer sequences for quantitative real-time PCR are summarized in Table 1.
Statistical analysis
Circadian gene expression levels in mRNA and protein at 24 hr after ECS were analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. Relative expression of circadian genes was analyzed using mixed-effect model repeated measure (MMRM) analysis, with six serial assessments over 24 hr (sampling times: ZT8, ZT12, ZT16, ZT20, ZT0, and ZT4) and two groups: sham vs. E1X, sham vs. E10X, and E1X vs. E10X. To test the effects of drugs or other stimuli on the expression of the genes showing circadian rhythms, it was reported to be reliable to apply the single cosinor method by fitting a 24 hr cosine curve to 6 values, 4 hrs apart [38]. The circadian rhythm of three variables was calculated using the single cosinor method to calculate the mesor (middle value of the fitted cosine representing a rhythm-adjusted mean), amplitude (half the difference between the maximum and minimum of the fitted cosine function), and acrophase (time of peak value in the fitted cosine function, expressed as the lag in hours and minutes from midnight) and standard error (SE) of their dispersion [39,40]. The six time-normalized means were also analyzed for a time effect by one-way ANOVA for the variables of circadian genes. The rhythm properties (mesor, amplitude, and acrophase) of each variable were compared between the groups by one-way ANOVA. All tests were performed using SPSS 19.0 for Windows (IBM Corp., Armonk, NY, USA). p-values <0.05 were deemed statistically significant.
RESULTS
mRNA level of circadian genes in the rat frontal cortex at 24 hr after ECS treatment
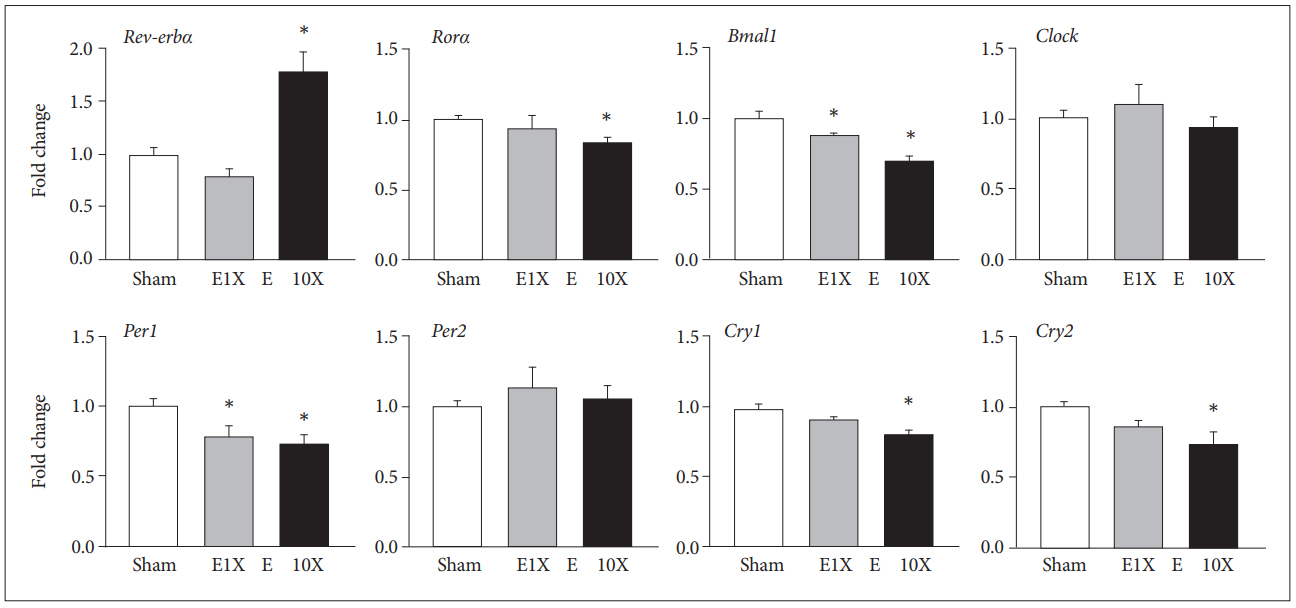
The mRNA level of circadian genes at 24 hr after single (E1X) or repeated (E10X) ECS treatment were examined. Compared with sham treatment, ECS treatment resulted in expression changes of Rev-erbα, Rorα, Bmal1, Per1, Cry1, and Cry2, whereas it showed no effect on the expression of Clock and Per2 (Figure 1). Rev-erbα expression was significantly changed (F=24.12, p=0.01) which was increased by E10X (p=0.01) but not by E1X. The expression levels of Rorα, Cry1, and Cry2 were significantly changed (F=3.59, p=0.03; F=10.47, p=0.02; F=4.77, p=0.03, respectively) which was significantly decreased by E10X (p=0.02, p=0.03, p=0.02, respectively) but not by E1X. Bmal1 and Per1 were also significantly affected by ECS treatments (F=27.61, p<0.01; F=5.94, p=0.03) decreased by both E1X and E10X (Bmal1 p=0.01, p=0.02; Per1 p=0.03, p=0.02) compared with sham.
Protein level of circadian genes at 24 hr after ECS treatment in the rat frontal cortex
E10X induced changes in protein levels in circadian genes, but E1X did not show any significant changes compared with sham. Rev-erbα was increased by E10X (F=3.60, p=0.03), whereas Rorα (F=3.55, p=0.03), Bmal1 (F=11.7, p=0.02), Clock (F=10.84, p=0.02), and Per1 (F=8.58, p=0.02) were decreased significantly compared with sham (Figure 2). However, the protein level of Cry1 was not changed by ECS treatment, and that of Cry2 showed an increase in E10X (F=5.44, p=0.03), whereas its mRNA level decreased (Figure 1).
Daily pattern of circadian gene expression in the frontal cortex
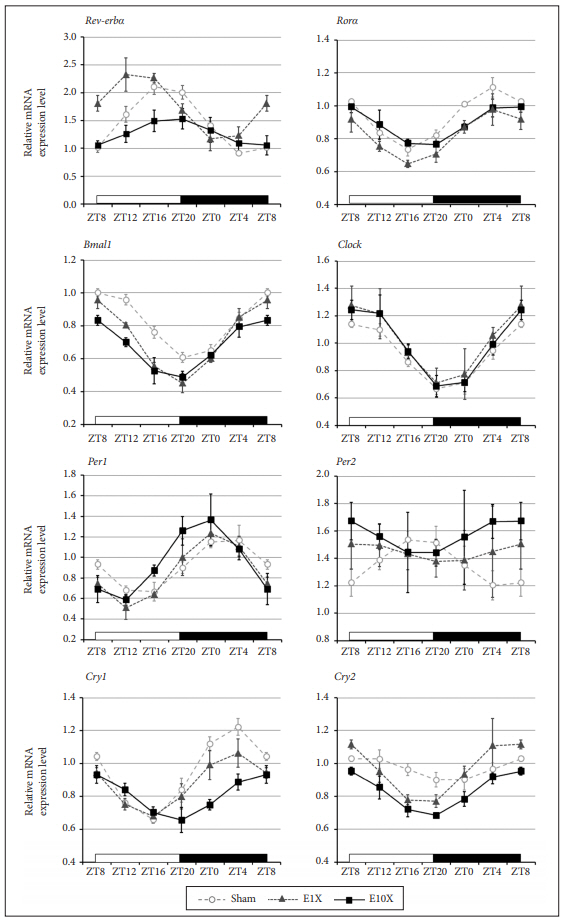
We investigated the effect of ECS on the daily oscillations of circadian gene expression. The effects of ECS on the daily expression of circadian genes were examined in the frontal cortex at 4, 8, 12, 18, 20, and 24 h after ECS treatments.
Rev-erbα
Rev-erbα expression showed a statistically significant daily rhythm in sham, E1X, and E10X (Table 2). E1X lowered Rev-erbα expression (F=6.49, p=0.03), and a group×time interaction was observed (F=3.73, p=0.04) compared with sham. E10X increased Rev-erbα expression (F=19.10, p=0.02), and a group ×time interaction was found (F=7.01, p<0.01) compared with sham. The mesor was significantly affected (F= 17.58, p=0.02) that decreased in E1X (p=0.03) and increased in E10X (p=0.01) compared with sham. The acrophase was observed approximately 3 hr 40 min earlier in E10X (p=0.02) than in sham (Figure 3, Table 2).
Rorα
Rorα expression showed a statistically significant daily rhythm in sham, E1X, and E10X (Table 2). Rorα expression level was not changed by E1X, while a group×time interaction (F=2.52, p=0.04) compared with sham was observed. E10X lowered Rorα expression (F=21.25, p=0.02), and no group×time interaction was observed. The mesor was decreased in E10X (F=7.64, p=0.02), but the acrophase and amplitude showed no changes after ECS treatment (Figure 3, Table 2).
Bmal1
Bmal1 expression showed a statistically significant daily rhythm in sham, E1X, and E10X (Table 2). Bmal1 expression was decreased by both E1X (F=44.69, p<0.01) and E10X (F=18.95, p<0.01), and group×time interactions were noted in E1X (F=2.49, p=0.03) and in E10X (F=3.61, p=0.02) compared with sham. The mesor was significantly affected (F=12.27, p=0.02) that decreased in E1X (p=0.03) and E10X (p=0.03) compared with sham. The acrophase was significantly affected (F=10.48, p=0.01) that approximately 1 hr 40 min earlier in E1X (p=0.03) and 2 hr 20 min earlier in E10X (p=0.02) than in sham (Figure 3, Table 2).
Per1
Per1 expression showed a statistically significant daily rhythm in sham, E1X, and E10X (Table 2). No significant difference in Per1 expression was observed in E1X and E10X compared with sham. The mesor and amplitude showed no difference in both E1X and E10X. The acrophase was observed approximately 3 hr 13 min earlier in E10X than in sham (F=8.31, p<0.01) (Figure 3, Table 2).
Per2
Per2 expression showed a statistically significant daily rhythm in sham but not in E1X or E10X (Table 2). There was no difference in Per2 expression between sham and E1X, while E10X increased Per2 expression (F=17.31, p<0.01) compared with sham. The mesor was increased in E10X (F=4.37, p=0.03), and the amplitude showed no difference compared with sham. The acrophase was observed 11 hr 28 min earlier in E10X than in sham (F=6.29, p=0.03) (Figure 3, Table 2).
Cry1
Cry1 expression showed a statistically significant daily rhythm in sham, E1X, and E10X (Table 2). Both E1X and E10X lowered Cry1 expression compared with sham (F=8.28, p=0.02; F=31.13, p<0.01). A group×time interaction was observed between sham and E10X (F=9.61, p<0.01). The mesor was significantly affected (F=18.76, p<0.01) that decreased in E1X (p=0.03) and E10X (p<0.01). The amplitude was also affected (F=11.49, p=0.01) that decreased in E1X (p=0.02) and E10X (p<0.01). The acrophase occurred approximately 3 hr 50 min later in E10X than in sham (F=16.63, p<0.01) (Figure 3, Table 2).
Cry2
Cry2 expression showed a statistically significant daily rhythm in sham, E1X, and E10X (Table 2). E10X decreased Cry2 expression compared with sham (F=31.44, p<0.01). The mesor was decreased in E10X (F=11.37, p<0.01) compared with sham. The acrophase was significantly changed (F=4.06, p=0.03) which occurred approximately 4 hr 25 min earlier in E1X (p=0.03) and 3 hr 35 min earlier in E10X (p=0.03) than in sham (Figure 3, Table 2).
DISCUSSION
We have demonstrated that ECS alters the expression and daily oscillation of circadian genes in the rat frontal cortex. The ECS-induced changes were more remarkable after repeated treatments than after single treatment suggesting the enhanced effects with the increasing number of ECS. Reduced mRNA expression of Rorα, Bmal1, Clock, Per1, and Cry1, and earlier acrophase of Rev-erbα, Bmal1, Per1, Per2, and Cry2 suggested the phase advance accompanied with attenuated transcription after repeated ECS treatments.
Circadian gene expression is regulated by integrated feedback loops [22]. Bmal1 and Clock dimerize and activate transcription of Per1, Per2, Cry1, and Cry2 through binding to the E-box on their promoter regions [41,42]. Rev-erbα suppresses Bmal1 expression via the RORE motif on its promoter region [43]. Rorα activates Bmal1 transcription by competing with Reverbα to bind to the RORE on the Bmal1 promoter [44]. Therefore, increased Rev-erbα along with reduced Rorα could result in the reduced Bmal1 which could relate to the decreased transcription of Per1, Cry1, and Cry2 after E10X treatment. However, the transcription of these genes are regulated by various mechanisms other than circadian genes themselves, which requires further investigations.
Another possible regulatory mechanism could relate to immediate early gene, Egr1. We have reported that repeated ECS treatments induced prolonged down-regulation of Egr1 expression in the rat frontal cortex [28,30]. Egr1 was reported to induce the expression of Rorα, Bmal1, and Per1, and to reduce that of Rev-erbα, Rev-erbβ and Per2 [45]. E10X-induced decrease in the expression of Rorα, Bmal1, and Per1 and increased Rev-erbα expression might relate to the reduced Egr1 level.
All the examined genes showed statistically significant daily oscillation in the frontal cortex of the sham-treated control rats. The present findings support the many previous reports that have demonstrated the diurnal rhythm of clock genes in the frontal cortex [19,46,47]. Mesor of Rev-erbα was increased, and that of Rorα, Bmal1, and Cry1 was reduced in response to E10X. Amplitude was not affected by ECS treatments. Among the examined eight genes, five genes, including Rev-erbα, Bmal1, Per1, Per2, and Cry2, showed earlier acrophase after E10X.
Our findings showing the phase advance in the expression of several circadian genes could be compatible with the effects of ECT and other antidepressants on circadian rhythm. The patients with depressive disorders who reached clinical remission after repeated ECT showed the advanced acrophase of the circadian rhythm [48]. Antidepressants, including agomelatine and trazodone, induce a phase advance [49,50]. Nonpharmacological treatment modalities for depressive disorder, such as sleep deprivation and light therapy, induce phase advance [51,52]. Antidepressants and mood stabilizers could reset abnormal clock genes in mood disorders in order to restore and stabilize circadian rhythm [11]. ECS-induced changes in circadian genes might contribute to the correction of mood instability.
The protein level showed similar pattern of changes compared with mRNA level after ECS except Cry2. mRNA level of Cry2 was decreased, whereas protein level was increased by E10X. Protein degradation through phoshporylation and ubiqutination is the strong contributing factor to the setting of circadian proteins level [23,53]. Phosphorylation of Cry2 at Ser 553/557 followed by ubiquitination leads to the proteasomal degradation [54,55]. Further studies will be necessary to understand the regulation of protein level of Cry2 by repeated ECS treatments.
The present study has several limitations. We performed experiments in normal animals, but not in animal models showing disturbances in circadian rhythm. Additional studies using disease animal models could enable us to understand the ECS’s effects on circadian genes related to the behaviors and multiple physiological variables [56]. Rhythm of circadian genes in various brain regions shows not only the phase synchrony across regions but also the region-specific differential patterns [19]. Studies on multiple brain regions, including SCN, could provide the differential and common effects of ECT on circadian genes across the brain regions. In addition, further studies will be necessary to understand the underlying molecular mechanisms inducing the changes in circadian genes by ECS treatemnts.
In conclusion, the present study shows that ECS, more prominently with repeated treatments, affects the expression and daily oscillation of circadian genes in the rat frontal cortex. Although each gene did not change in a uniform way, reduced transcription and phase advance of several genes after repeated ECS treatments were remarkable. ECT-induced changes in circadian genes could relate to the therapeutic effects, which might provide the potential molecular targets for the treatment of mood disorders.