INTRODUCTION
Panic disorder (PD)-encompassing psychological, neurological, and biological dimensions-is an anxiety disorder characterized by predictive anxiety, unexpected panic attacks, and physical symptoms. These symptoms include sensations of shortness of breath, palpitation, or fear of dying. Recently, research on neurobiological and genetic factors of PD has been emerged. Previously, several studies suggested that genetic factors might contribute to oneâs susceptibility of developing PD. The relative risk in first-degree relatives of proband with PD between 2.6 to 20 times higher [1]. A number of twin studies have further demonstrated that over 40% of the identified liability toward PD can explain the heritable factors thereof [2,3].
The brain-derived neurotrophic factor (BDNF) protein is encoded by a gene found in humans on chromosome 11 and is part of the neurotrophin family of nerve growth factor essential for neurodevelopment. In the brain, the BDNF effects the hippocampus, cortex, and basal forebrain-areas vital to memory, learning, and higher thinking [4]. Single nucleotide polymorphisms (SNPs) of BDNF have been widely investigated across various psychiatry disorders, and there are significant associations between BDNF polymorphisms and susceptibility of schizophrenia [5], major depressive disorders [6], and Alzheimerâs disease [7]. In particular, rs6265 which produces an amino acid substitution from valine (Val) to methionine (Met) in the BDNF gene [8] is the most investigated SNP throughout the field of psychiatric [9].
A previous study (of non-clinical samples) suggests that the GG (Val/Val) genotype of rs6265 in BDNF show increased trait anxiety as measured via the State Trait Anxiety Inventory compared to those of the GA (Val/Met) genotype [10]. A meta-analysis of patients with anxiety disorders show that the rs6265 GG genotype results in increased neuroticism scale scores compared to A-carriers [11]. Although there is a lack of previous findings regarding the association of rs6265 with anxiety-related traits in patients with PD, some studies suggest that BDNF rs6265, in particular, plays a role in psychopathology-such as neuroticism or trait anxiety-in PD [12,13]. A further study investigates the association between rs6265 and trait anxiety in individuals with PD, showing lower trait anxiety scores in those with the GG genotype of rs6265 than individuals of the GA/AA genotype in the early onset PD group [13]. The other study suggests that BDNF SNP rs6265 had an interaction effect with Catechol-O-methyltransferase gene (COMT) polymorphism to neuroticism scores and anxiety levels in patients with PD [12].
So far, the amygdala, hippocampus, frontal cortex, and sensory cortex areas-as they relate to the fear network model of PD-has been studied extensively [14]. Recently, however, it has been suggested that the white matter (WM) microalterations of the corpus callosum (CC) might be associated with visual memory-related processing and cognitive and affective functions [15]-functions that may be major components of PD patients with higher anxiety sensitivity in their visual dependence and in perceiving external threatening stressors [16]. The CC is the largest WM tract in the brain, integrating information between the hemispheres so as to control recognition and emotional functions [17-19], and is specifically associated with attention, anxiety, and depressive symptoms [20]. However, there are a limited number of studies on the association between the CC and pathophysiology, anxiety-related traits, and symptoms in patients with PD.
Further research investigating the association between the BDNF rs6265 polymorphism and brain structural abnormalities (including WM) has also emerged. Previous studies show that the BDNF rs6265 is associated with a reduction in hippocampal tissue (a key structure for the encoding of emotionally relevant data into memory) as well as with anxiety disorders [21] and major depressive disorders [22]. Besides the genetic knowledge of limbic structures, a recent study showed that the A-carriers of rs6265 in healthy adults have higher structural WM connectivity in the CC [23]. Interestingly, the association between BDNF rs6265 and WM lesions of the frontal CC is present in elderly patients with major depressive disorder [24]. Thus, we considered the need for further research on the association between the BDNF rs6265 polymorphism and the WM connectivity of the CC in PD.
The aim of our study is to determine 1) whether the BDNF Val66Met polymorphism is associated with the WM connectivity of the CC in PD and 2) whether it is correlated with anxiety-related trait vulnerability and measures of symptom severities. We formulated the following hypotheses: 1) there would be differences of the WM connectivity in the CC between the GG genotype group and the non-GG genotype group; 2) that there would be gaps of trait vulnerability and symptom severities scale scores between the two groups; and 3) that WM connectivity in the CC would be associated with trait vulnerability and symptom severity measures.
METHODS
Participants
A total of 95 patients with PD participated in this study. Between January 2011 and May 2019, study participants were recruited from the pool of PD patients treated in the Department of Psychiatry at CHA Bundang Medical Center (Gyeonggi-do, Korea). All participants were between 17 and 64 years of age, right-handed, and of Korean descent. Individual and family histories of PD patients were collected through interviews.
Participants with PD-with or without agoraphobia-were diagnosed by practiced psychiatrists using the Structured Clinical Interview to evaluate DSM-VI-TR (as per the Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition, Text Revision (DSM-IV-TR) Axis I Disorder). Only patients with primary PD were included. Secondary lifetime diagnoses included major depressive disorders (17 patients) and generalized anxiety disorder (10 patients). Exclusion criteria included any history of schizophrenia, intellectual disabilities, alcohol and substance abuse or dependence, and current or past serious medical disorders-including neurological disorders, pregnancy, and contraindications to brain magnetic resonance imaging (MRI) scanning.
At the time of MRI acquisition, 78 PD patients were undergoing pharmacotherapy with selective serotonin reuptake inhibitors (SSRIs), including escitalopram, paroxetine, and sertraline [n=78; escitalopram equivalence dosage, mean (ÂąSD) 9.33Âą7.48 mg/day], and benzodiazepines as anxiolytics, including lorazepam, alprazolam, clonazepam and diazepam [n=88; lorazepam equivalence dosage mean (ÂąSD) 1.92Âą1.96 mg/day]. Brain MRIs were performed 11.55Âą24.11 days after the initiation of medication uptake. All patients with PD were undergoing pharmacotherapy with antidepressants and anxiolytics according to the Korean Mediation Algorithm for PD [25] or Clinical Practice Guidelines: Treatment of PD [26].
All study procedures conform to the Institutional Review Board regulations and the principles of Good Clinical Practice delineated by the CHA Bundang Medical Center. After a sufficient explanation of the study was given to the participants, written informed consent was obtained from each participant (2019-05-030, 2018-06-029, 2011-11-164).
Clinical assessments
The Neuroticism-Extraversion-Openness (NEO)-neuroticism Inventory [27] and the Korean version of the Anxiety Sensitivity Inventory-Revised (ASI-R) [28,29] measurements were used at an early stage to evaluate potential trait markers for PD. The NEO-neuroticism inventory contains a total of 12 items and has a high internal consistency, ranging from 0.68 to 0.86 [30]. Meanwhile, the ASI-R is the most commonly-used measure of sensitivity to anxiety and consists of the following items: fear of respiratory symptom; fear of cardiovascular symptom; fear of a publicly observable anxiety reaction; and fear of cognitive dyscontrol. The internal consistency coefficient of the Korean version of the ASI-R is 0.92 and its test-retest reliability is measured to be 0.82.
Genotyping
To analyze the BDNF Val66Met (rs6265) polymorphism, Genomic DNA was extracted from peripheral whole blood samples using a G-DEXTM II Genomic DNA Extraction Kit per manufacturerâs protocol (Intron Biotechnology, Korea). The detection of functional SNPs in the genes encoding BDNF was based on an analysis of primer extension products as generated out of the genomic DNA, and which has been amplified using a chip-based matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry platform (Sequenom Inc., San Diego, CA, USA). The primer sequence for BDNF Val66Met was 5'-ACG TTG GAT GCT TCA TTG GGC CGA ACT TTC-3'. The general procedures were performed according to the manufacturerâs standard protocol [37]. Details of the genotyping procedures have been described above.
The genotype frequencies of BDNF Val66Met for Asian populations have been reported as follows [38]: Korean, AA (Met/Met) genotype 23%, GA (Val/Met) genotype 46%, GG (Val/Val) genotype 31%. We grouped PD patients into GG and non-GG groups, the reasons are as follows: first, the BDNF Val66Met polymorphism genotypes were grouped across Met/Met and Met/Val in accordance with much of the literature, as the homozygous Met/Met genotype is relatively smaller across populations; Secondly, previous studies show well-validated genotype grouping along GG and non-GG groups [10,39,40].
Image acquisition
Magnetic resonance images were acquired using a 3 Tesla scanner (SignaHDxt, GE Healthcare, Milwaukee, WI, USA). Diffusion-weighted imaging (DWI) [41] was performed using an echo planar imaging (EPI) sequence using the following parameters: repetition time (17,000 ms), echo time (108 ms), field of view (240 mm), acquisition matrix (144Ă144), slice thickness (1.7 mm), and voxel size (1.67Ă1.67Ă1.7 mm) [3,42]. We use a double-echo option to decrease eddy current-related distortions. To reduce the effects of EPI spatial distortions, we made use of an 8-channel coil and array of spatial sensitivity encoding (SENSE), sped-up by factor of 2. Seventy axial slices-parallel to the anterior commissure-posterior commissure line encompassing the entire brain-were obtained in fifty-one directions with b=900 s/mm2. Eight baseline scans with b0=0 s/mm2 were performed. Diffusion tensor images (DTIs) were measured from DWI utilizing the least-squares method (approximate scan time of 17 minutes).
Tract-based spatial statistics
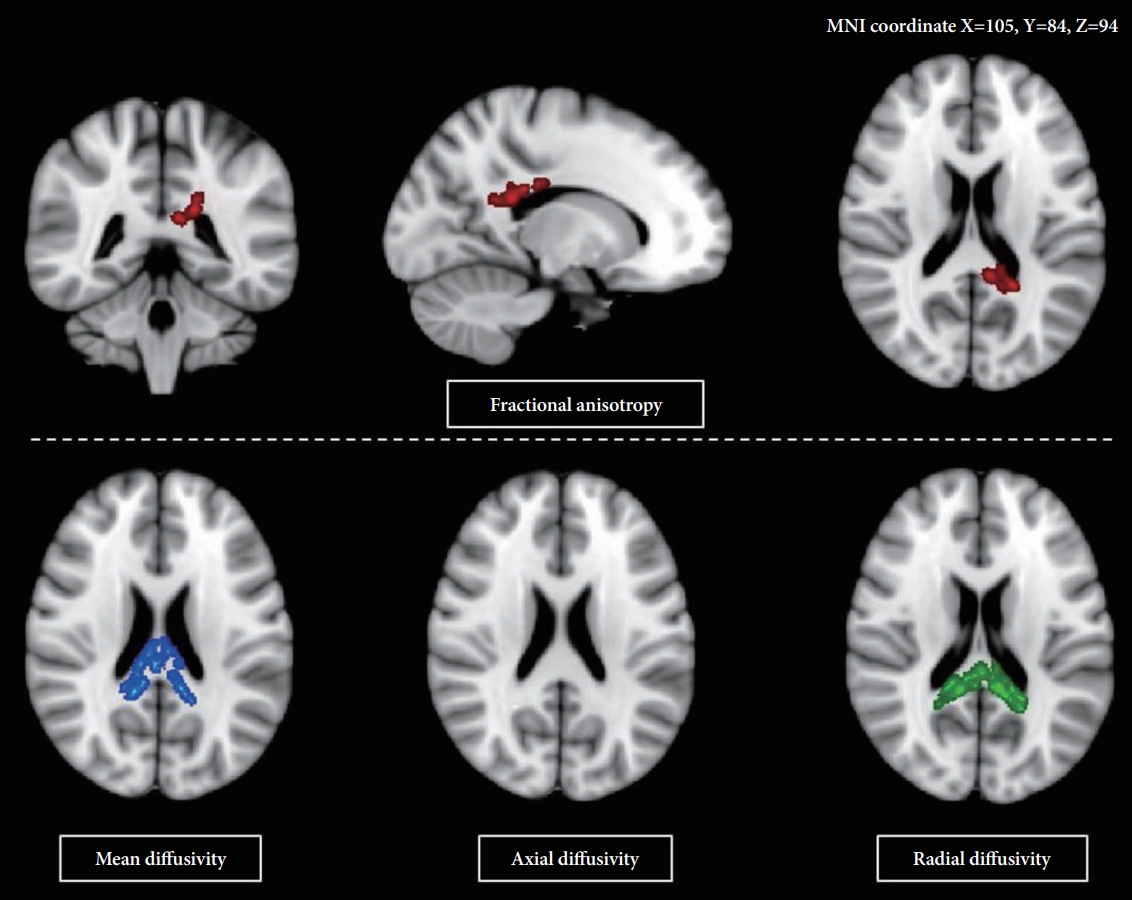
Voxelwise statistical analysis about fractional anisotropy (FA) data was performed using Tract-Based Spatial Statistics (TBSS) version 1.2, as implemented via the Functional Magnetic Resonance Imaging of the Brain (FMRIB) Software Library (FSL version 4.1, Oxford, UK, https://fsl.fmrib.ox.ac.uk/fsl) according to the standard procedure [43]. The FA is a measure of the extent of an ellipsoid, and provides information about the degree of anisotropy in the voxel. First, the DTI preprocessing-which included skull stripping using the Brain Extraction Tool and eddy current correction-was performed using the FMRIB Software Library (FSL) [44]. We used the FMRIBâs Nonlinear Image Registration Tool to align all subjectsâ FA data into the standard space (Montreal Neurologic Institute 152 standard). All transformed FA images were then integrated and applied to the original FA map so as to create a standard-space version thereof. All transformed FA images were averaged to produce a mean FA image, which was then skeletonized to make a mean FA skeleton which only considered the center of the WM tracts. The threshold of the skeleton was set to FA>0.2 (the TBSS default) so as to only contain major fiber bundles.
To compare DTI indices [FA, mean diffusivity (MD), axial diffusivity (AD), and radial diffusivity (RD)], FSL FA images were used in order to achieve nonlinear registration and obtain stages of skeletonization, and to estimate the projection vectors from each individual subject onto the mean FA skeleton [45]. The nonlinear warps and skeleton projection were applied to other images.
Mean FA skeletons were multiplied using the Johns Hopkins University DTI-based probabilistic tractography atlas, containing the CC [46].
Statistical analysis
We performed a voxel by voxel statistical analysis in order to detect regions of significant differences in the scores of the DTI indices (i.e., FA, MD, AD, and RD) among genotype groups [the BDNF GG genotype group vs. non-GG (GA/AA) genotype group], respectively. We conducted this analysis using nonparametric permutation tests with a correction for multiple comparisons with the FSL Randomize program. In addition, analysis of covariance (ANCOVA) and multiple linear regression analysis were conducted making use of age, sex, intracranial volume (ICV), neuroticism, and BDI-II as covariates. This allowed us to confirm the effect of other variables upon the results.
To confirm the voxel-wise analysis results, the DTI indices of the CC were extracted; the indices demonstrated significant differences in the FA, MD, and RD values between the two groups. These results were applied to further analysis. We conducted correlation analyses to investigate whether regional differences in the FA value could potentially be associated with the variance in clinical symptom ratings among in each genotype group.
We performed statistical analyses using SPSS version 26.0 (IBM Corporation, Armonk, NY, USA). Differences with p<0.05 were statistically significant.
RESULTS
Genotyping
We divided the participants with PD into either a GG genotype or a non-GG genotype (A-allele carrier) groups of the BDNF Val66Met polymorphism, in order to evaluate the differences between the groupsâ WM connectivity in their CCs.
The genotype distributions of all 95 patients with PD (GG: n=25, GA: n=53, AA: n=17) were in accordance with the Hardy-Weinberg equilibrium (Ď2=1.46, df=1, p=0.23). Upon division of the participants with PD into the two aforementioned groups, they were not significantly different in terms of sex (p=0.11) or age (p=0.72) (Table 1).
Comparisons of trait vulnerability and symptom severity measures between the GG genotype group and non-GG genotype group with panic disorder
The GG genotype group showed significantly higher scores the NEO-neuroticism, and BDI-II scales. However, there was no significant difference via the ASI-R and its subscales (Table 2).
Comparison of the diffusion tensor imaging indices of the corpus callosum between the GG genotype group and non-GG genotype group with panic disorder
Increased FA values in the posterior part of body and splenium of the CC were found in participants with the GG genotype compared to the non-GG genotype participants. However, the MD and RD values showed inverse results from the FA values (Figure 1). Significance remained the same after controlling for age, sex, the presence of comorbidity, and intracranial volumes.
Linear regression of BDNF Val66Met polymorphism on the fractional anisotropy values of body and splenium of the corpus callosum with panic disorder
Although the age and sex of PD patients did not show any significant correlation or association with the FA of the body and splenium of the CC, we decided to include age and sex so as to control for the potential confounding factors apparent in the regression model. Results of the linear regression of the FA values of the body and splenium of the CC on BDNF Val66Met genotype in patients with PD are presented in Table 3. This analysis reveals an association of BDNF Val66Met genotype and the FA values of the body (β=-0.43, p<0.001) and splenium of the CC (β=-0.60, p<0.001). Specifically, the FA values of the body and splenium of the CC in the GG genotype group were higher than those in the non-GG genotype group after controlling for age, sex, neuroticism, and BDI-II scores.
Exploratory correlation analysis of the diffusion tensor imaging indices of the corpus callosum and trait vulnerability and clinical symptom severity scores in the Val66Met genotype groups with panic disorder
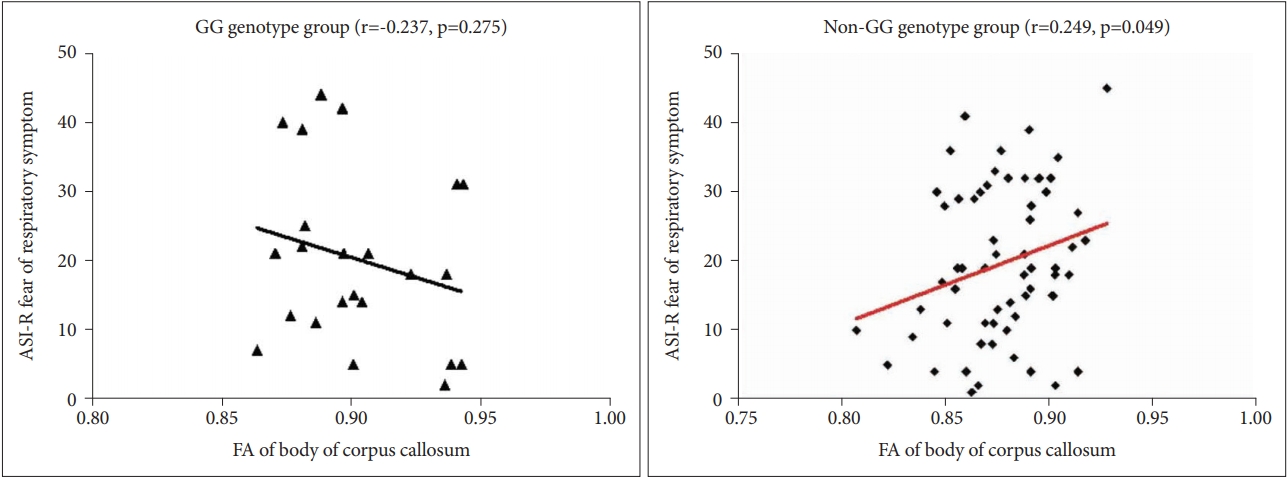
Correlation analysis between the WM regions in the body and splenium of the CC-where the FA values significantly differed, according to genotype between the two groupsâ trait vulnerability scores-reveals significant positive correlations between the FA values of the body of the CC regions, as well as the baseline ASI-R âfear of respiratory symptomâ scores. In the GG genotype group, a positive and statistically significant correlation is shown, unlike in the non-GG genotype group (r=0.249, p=0.049) (Figure 2).
DISCUSSION
To our knowledge, this is the first study to demonstrate that in patients with PD 1) the GG genotype group has significantly higher FA values in regard to the posterior part of the body and splenium of the CC, and 2) that the GG genotype group exhibits higher degree of neuroticism-a vulnerability trait of PD-and depressive symptoms than patients in the non-GG genotype group. Furthermore, among the non-GG genotype group, PD patients with higher FA values in the body of the CC showed higher baseline ASI-R âfear of respiratory symptomâ subscale scores. Our findings suggest that BDNF Val66Met polymorphism is associated with WM connectivity of the body and splenium of the CC, possibly associated with neuroticism and depressive symptoms in patients with PD.
In the present study, we observe that measures of neuroticism are higher among patients in the GG genotype group. Our results are similar to those of a the previous study of heterogeneous anxiety disorders, that included PD, post-traumatic stress disorder, obsessive-compulsive disorder, phobic disorder, and generalized anxiety disorder [11]. This similarity is evident in the observation that patients with PD in the GG genotype group display greater levels of neuroticism than those patients in the non-GG genotype group [11]. Moreover, since neuroticism can be associated with depressive symptomatology, we also find more depressive symptoms in patients of the GG genotype group rather than those in the non-GG genotype group.
In this study, we further find that patients in the GG genotype group show higher FA values in the body and splenium areas of the CC, as well as lower MD and RD values when compared to those in the non-GG genotype group. These findings suggest the possibility of a relationship between axon and myelin microalterations in the body and splenium regions of the CC of PD patients, according to the BDNF Val66Met polymorphism genotype. This may be evidence of WM abnormalities in PD patients, suggesting that microalterations in the WM contribute to the development of neuroticism and depressive symptoms in these patients.
The reason why the BDNF Val66Met polymorphism genotype is associated with the body and splenium areas of the CC is unknown. However, it has been suggested that the splenium of the CC can be considered a neocortical component of the hippocampal commissure, carriers fibers connecting the hippocampi with those linking the posterior parietal, medial temporal, and medial occipital cortices of the two hemispheres [47-49]. In addition, in emotional disorders (e.g., depressive disorder), the BDNF Val66Met polymorphism is related to the WM connectivity of the CC, including the body and splenium regions thereof [50]. As depressive disorder can be comorbid with PD, and as PD can share the depressive symptomatology with depressive disorder as well, it is likely that the BDNF Val66Met polymorphism likely can affect the splenium of the CC connectivity in those with PD. However, our results as derived by multiple linear regressions show that the FA values of the body and splenium of the CC are higher than in those patients of the non-GG genotype group when controlling for age, sex, BDI-II, and neuroticism. This result supports our hypotheses that BDNF Val66Met polymorphism can play an important role in explaining the microalterations of the body and splenium of the CC in PD patients regardless of age, sex, BDI-II, and neuroticism.
Literature regarding genetic-neuroimaging on BDNF has investigated whether the Val66Met polymorphism influences WM abnormalities in healthy adults [51,52]. In particular, previous results on BDNF Val66Met polymorphism in healthy adults present lower FA values in patients in the GG genotype group, contrary to our results of patients with PD [52]. We explain this inconsistency as follows: first, certain studies suggest the microalterations of the CCâs WM show higher FA values in patients with anxiety disorders, including PD [53-55]. The second reason for these is unclear but may relate to differences in sampling and biotype grouping. Additionally, our sample was limited to Korean individuals, and certain previous studies which only include Caucasian individuals [52].
Our study also finds a positive association between the ASIR âfear of respiratory symptomâ subscale score and the FA values in the body of the CC among patients in the non-GG genotype group. This result suggests that the WM connectivity of the body of the CC among PD patients in the non-GG genotype group might contribute to the trait vulnerability of anxiety sensitivity.
There were a number of limitations to our study. First, the sample size in each group was relatively small. Although significantly increased FA values in the CC regions were found among patients in the GG genotype group compared to the non-GG genotype group, further studies should include a larger sample for more accurate results. Secondly, only patients with PD were studied. To obtain the further understandings of the relationship between the BDNF Val66Met polymorphism and the CC of WM connectivity (such as genotype by diagnosis interaction), further studies should include a healthy control group. Third, the influence of medication on the WM connectivity in the CC across genotype groups could not be ruled out completely although the mean duration of medication day is relatively short.
In conclusion, the current findings suggest that the BDNF Val66Met polymorphism is associated with WM connectivity of the body and splenium of the CC, and that it may also be related to neuroticism and depressive symptoms in individuals with PD. Furthermore, PD patients in the non-GG genotype group, those with higher FA values in the body of the CC show higher anxiety sensitivity. These results demonstrate that the CC microalterations (according to the BDNF Val66Met polymorphism) are significant in the psychopathology of PD as depressive disorders.