Effects of 5HT1A Activation on Depression Profile Following 5-HT Depletion in Rats Lacking Social Attachment Since Weanling
Article information
Abstract
Objective
Post weanling isolation-reared (IR) rats are featured with depressive phenotype, yet its mechanism is not clearly defined particularly in terms of the involvement of central 5-HT1A receptors. The present study aims to examine the effects of 5HT1A activation on forced swim test (FST) in IR rats following 5-HT depletion.
Methods
Social control (SOC) and IR rats received an intracerebraoventricular (ICV) injection of 5-HT depletion agent, 5,7-DHT. 14 days after the surgery, rats were assessed their performance in FST with or without the challenge with a 5-HT1A agonist, 8-OH-DPAT. Rats were then sacrificed for analyzing their 5-HT tissue levels and the expressions of their 5-HA1A receptors in prefrontal cortex (PFC), hippocampus (HPX), and amygdala (AMY).
Results
5,7-DHT decreased the tissue concentration of 5-HT in both IR and SOC rats. IR rats were more immobile and less sensitive to the lesion-induced immobility, however this effect was reversed by acute challenge of 8-OH-DPAT. 5,7-DHT lesion increased the expression of PFC 5-HT1A receptors.
Conclusion
The integrity of central 5-HT system is developmentally crucial for the 5-HT1A-relevant depression profile in rats of social isolation.
INTRODUCTION
Rearing animals in a situation of social isolation since early life has profound effects on their psychological, behavioral, and neurochemical profiles [1-6]. One of the hallmark findings is that post weanling isolation-reared (IR) rats may exhibit depressive phenotype in their adulthood. For example, IR rats were found more immobile than their social controls in forced swim test (FST, an acknowledged depression paradigm) [7], indicating a behavioral despair in which the animal gives up to struggle out of a confined distressing situation [8]. However, the IR effects on the depression profile subject to different central nervous systems are not fully understood and it is worth to clarify.
Central serotonergic (5-HT) system is involved in the pathoetiology of depression, yet its precise role in the depression profile is not clearly defined. Two methods are usually employed to approach this issue, to activate the 5-HT system by using 8-OH-DPAT, a 5-HT1A receptor agonist; and on the contrary to lesion the presynaptic 5-HT with 5-HT depletion method by 5,7-dihydroxytryptoamine, 5,7-DHT. Studies of 5-HT activation in the development-associated depression are inconsistent. For example, intervention of 8-OH-DPAT at 0.5 mg/kg successively reversed the increased immobility time of FST in a rodent model of chronic pain induced depression [9]. On the other hand, if the 5-HT autoreceptor was knockdown, the duration of immobility was shortened too [10]. This discrepancy in fact highlights the different involvements of presynaptic effect of dorsal raphe nucleus and post-synaptic effect in hippocampus in the depression profile [11]. It is particularly relevant to the depletion method as mentioned above as it specifically removes the contributive role of 5-HT1A somatodendritic autoreceptor which normally is highly relevant to the presynaptic or releasing performance of 5-HT.
Increasing evidence reveals that manipulation of the social experience alters stress-related neural circuits and monoaminergic systems [12], in which the procedure of IR exerts considerable impact on the central 5-HT function. We previously demonstrated that for IR rats, 8-OH-DPAT strengthened prepulse inhibition (PPI, an ability of sensorimotor gating) in sham rats but downgraded it in depletion condition [13], suggesting a possibility of multiple roles of 5-HT1A receptor in the IR-associated depression profile. In this regards, it is worth investigating whether 5-HT depletion model, as it downgrades the presynaptic 5-HT effects, could be used to explore the role of 5-HT1A receptor on FST performance in rats of ongoing social deprivation since weanling, which is particularly necessary in exploring the mechanism underlying the 5-HT1A modulation onto the development-associated depression profile. In the present study, social control (SOC) and IR rats received an intracerebraoventricular (ICV) injection of 5-HT depletion agent, 5,7-DHT. With 14 days recovery from the surgery, rats were assessed their performance in FST with or without the challenge of a 5-HT1A agonist, 8-OH-DPAT. At the end of the study, animals were sacrificed for analyzing their 5-HT tissue levels and the expressions of their 5-HA1A receptors in prefrontal cortex (PFC), hippocampus (HPX), and amygdala (AMY). Our results showed the integrity of central 5-HT system is developmentally crucial for the 5-HT1A-relevant depression profile in rats of social isolation. It may be helpful for a better understanding in terms of the functional role of 5-HT1A receptor in developmentally social attachment.
Methods
Animals
Male Sprague-Dawley (SD) rats aged at 21–23 days old (weaned; BioLASCO Taiwan Co., Ltd., Yilan, Taiwan) were used in the study and were divided randomly into 2 rearing conditions [social rearing (SOC, 2 rats per cage, n=10) vs. isolation rearing (IR, housed singly, n=10)]. The animals were kept in a temperature-(22±4°C) and humidity-controlled (50±20%) room under a 12-h light/dark cycle (lights on from 07:00 to 19:00) and given ad libitum access to a standard laboratory chow diet (Ralston Purina, St. Louis, MO, USA) and sterile water. At their age of postnatal days 77–79, rats in each rearing condition were randomly assigned to one of the two managements (sham control vs. 5-HT depletion surgery, n=5 for each group), thus made it totally 4 groups (SOC-sham, IR-sham, SOC-depletion, IR-depletion, n=5 for each) to enter the FST protocol (14 days after the lesion surgery) for measuring their performance under 8-OH-DPAT. Tissue levels of 5-HT/5-HIAA and the protein expressions of 5-HT1A receptors in PFC, HPX, and AMY were also examined in sham and lesioned rats under different rearing conditions. The study was approved by the Institutional Animal Care and Use Committee of National Defense Medical Center with the certificate number NDMC-10319.
Locomotor activity
Total travel distance was used to reflect the locomotor activity, and was measured using a computerized automated activity monitoring system (MED Associates, Inc., St. Albans, VT, USA). The system included four plexiglass chambers (43×43×30 cm) equipped with an infrared array of 16 photodetectors and corresponding light sources that emitted photobeams 3 cm apart and 4.5 cm above the chamber floor. Travel distance was recorded every 5 min and expressed as cumulative 60-min data [13]. Locomotor activity was tested 2 days before FST.
Forced-swimming test
Forced-swimming test (FST) was performed as described previously by our team [14,15], and was performed between 9:30 A.M. and 1:00 P.M. In brief, the rats were placed for 15 min into a 25 cm diameter × 50 cm height plastic cylinder, which was filled with 20–25°C water to a depth of 30 cm. The rats were then removed, dried, and returned to their home cage. They were placed again in the cylinders 24 h later, and then a 5-min swim test was conducted and videotaped. Immobility was defined as the minimum movement required to passively keep the animal’s head above the water without other motions. Climbing was defined as the upward-directed movement of the forepaws against the wall. The results are expressed as the amount of time (in seconds) that the animals spent immobile and climbing during the 5-min test.
Surgery
Microinjection needle (Hamilton, 30-gauge, 10 μL) was used to inject 5,7-DHT into ICV (AP: -0.8 mm from bregma, L: ±1.5 mm from the midline, DV: -0.4 mm from dura) [16], with a flow rate of 1 μL/min controlled by a microinjection pump (CMA/100, Carnegie, Medicin, Sweden) for 10 min.
Drugs
Serotoninergic neurotoxin 5,7-DHT (200 μg/rat, Sigma‐Aldrich, St. Louis, MO, USA) was dissolved in saline solution with 0.6% ascorbic acid and was administered via intracerebraoventricular (ICV) injection. 30 min before the 5,7-DHT injection, desipramine (25 mg/kg, Sigma‐Aldrich), a reuptake inhibitor of norepinephrine, was administered intraperitoneally to protect noradrenergic neurons from neurotoxicity [17]. To activate the 5-HT1A receptors, 8-OH-DPAT (Sigma‐Aldrich) was intraperitoneally injected (1 mg/kg, in a volume of 1 mL/kg) 10 min prior to the behavioral testing [18].
Tissue levels of 5-HT and 5-HIAA in PFC, HPX, and AMY
Rats were sacrificed by decapitation and their brains were rapidly removed. Various brain areas, including prefrontal cortex, hippocampus, and amygdala were rapidly dissected on an icy cold plate, then weighed and stored at -80°C until homogenization with ultrasonication in 0.2 mL of 7N perchloric acid (Sigma Chemical Industries, Ltd., Saint Louis, MO, USA). Homogenates were centrifuged at 12,000×g for 30 min at 4°C. The concentration of serotonin (5-HT) and its metabolite 5-HIAA was determined by high performance liquid chromatography (HPLC) equipped with electrochemical detector (ECD, LC-4C, BAS, West Lafayette, IN, USA) (10 nA, filter 2.0 Hz, AppE cell 0.750 V), autosampler (Shimadzu SIL-10ADvp autosampler, Shimadzu, Japan). The supernatant was filtered through a 0.22 um filter, and was analyzed by HPLC equipped with an AlltimaTM (Grace Davison Discovery Scienes, Ltd., Deerfield, IL, USA) reversed phase C18 column (4.6×150 mm, 5 μm). Injection volume was 20 uL. The mobile phase contains 100 mM NaH2PO4.H2O, 0.74 mM sodium octanesulfonate (SOS), 0.02 mM EDTA and 20% methanol adjusted the pH to 3.0 by H3PO4 and with a flow rate of 1.0 mL/min. The calibration curve was obtained based on 6 levels (20, 40, 60, 80, 100, and 200 ppm) of standard of 5-HT and 5-HIAA (Sigma Chemical Industries, Ltd.). The coefficient of determination (r2) was greater than 0.995.
Western blotting
Equal amounts of brain tissue (100 μg) were denatured by heating at 95°C for 10 min, then separated by SDS-polyacrylamide gel electrophoresis using 10% polyacrylamide gels. Separated proteins in the gel were then electroblotted on to polyvinylidene difluoride membranes (Bio-Rad, Berkeley, CA, USA) for western blot analysis. Membranes were blocked with 5% bovine serum albumin in TBST (136.8 mM NaCl, 2.68 mM KCl, 24.7 mM Tris base, 0.1% Tween 20, pH: 7.4) at room temperature for 1 hour. The primary antibody 5-HT1A (Cat. No.: sc-1459, Santa Cruz Biotechnology, TX, USA) was used at 1:1,000–10,000 dilutions for incubation with membranes overnight at 4°C. Additionally, β-actin (Sigma; Cat. No.:A5441A) was used at a 1:500,000 dilution. After primary antibody incubation, the membranes were incubated with the corresponding secondary antibodies (1:5000 dilution) for 1 h at room temperature. Blots were then washed, and immunoreactive bands were detected by Immobilon™ Western Chemiluminescencent HRP substrate (Millipore; MA, USA Cat. No.: WBKLS0500) and recorded using a FUJI Medical X-ray film (Super RX-N; Cat. No.: 4741019291; Tokyo, Japan).
Data analyses
Statistical analyses were performed across the groups via a multi-factor analysis of variance (ANOVA) by using SPSS Version 18.0 (SPSS Inc., Chicago, IL, USA), with REARING CONDITION and LESION as between-subject factors. Further analyses with post-hoc multiple comparisons were performed where possible. All data were expressed as mean±standard error of the mean (SEM). A p value of <0.05 was considered statistically significant.
Results
Effects of 5,7-DHT on tissue levels of 5-HT and 5-HIAA
For demonstrating the lesion effects of 5,7-DHT, tissue levels of 5-HT and 5-HIAA of lesion group were compared with sham group in percentage. In SOC rats, concentration (mean, in pg/mg) of 5-HT/5-HIAA of sham groups were 651/7103 (PFC), 1366/14910 (HPX), and 537/6428 (AMY). HPLC method confirmed the reductions of tissue levels of 5-HT/5-HIAA caused by 5,7-DHT in PFC (5-HT: 46%; 5-HIAA: 56%), HPX (5-HT: 18%; 5-HIAA: 34%), and AMY (5-HT: 27%; 5-HIAA: 38%). In IR rats, concentration (mean, in pg/mg) of 5-HT/5-HIAA of sham groups were 480/5332 (PFC), 8673/10905 (HPX), and 339/2820 (AMY). HPLC method confirmed the reductions of tissue levels of 5-HT/5-HIAA caused by 5,7-DHT in PFC (5-HT: 22%; 5-HIAA: 28%), HPX (5-HT: 11%; 5-HIAA: 23%), and AMY (5-HT: 16%; 5-HIAA: 14%). For statistics, there were a significant main effect of LESION in PFC [F(1,16)=24.7, p<0.001], HPX [F(1,16)=77.7, p<0.001], and AMY [F(1,16)=55.9, p<0.001].
Locomotor activity
IR rats exhibited greater locomotor activity (indexed by total travel distance) in the 60 min activity test [REARING CONDITION, F(1,16)=5.60, p<0.05]. The treatment of 8-OH-DPAT did not cause any effect on locomotor activity in SOC rats, yet decreased the activity in IR rats under lesioned condition [F(1,16)=8.94, p<0.01] (Figure 1).
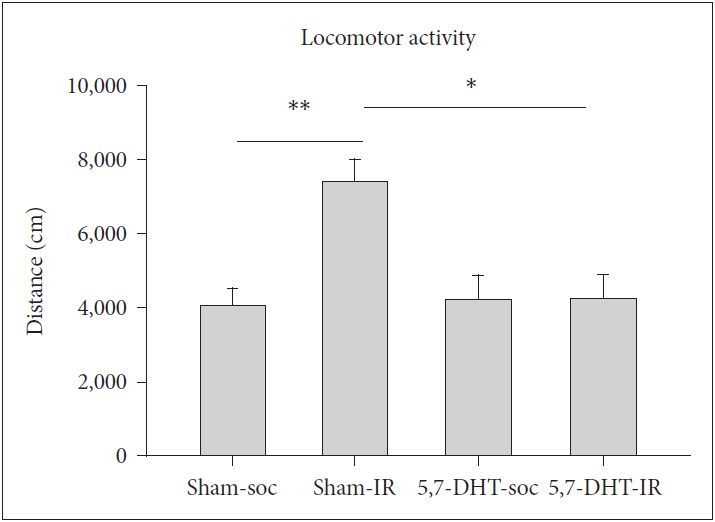
Effects of 8-OH-DPAT on locomotor activity in rats of IR and social control (SOC) (N=5, for each group). Scored as total travel distance in the 60 min activity test. Data are represented by mean±SEM. *p<0.05, **p<0.01. SEM: standard error of the mean.
Effects of 5,7-DHT on the performance of FST
There was a main effect of REARING CONDITION that IR rats exhibited greater score of immobility [F(1,16)=9.13, p<0.01] and lower score of swimming [F(1,16)=10.45, p<0.01] (Figure 2). For immobility, LESION caused a diverse effect, IR rats were less immobile whereas SOC rats became more immobile [REARING CONDITION×LESION, F(1,16)=8.72, p<0.01]. For swimming, LESION decrease the score in SOC but not IR rats [REARING CONDITION×LESION, F(1,16)=10.66, p<0.01] (Figure 2).
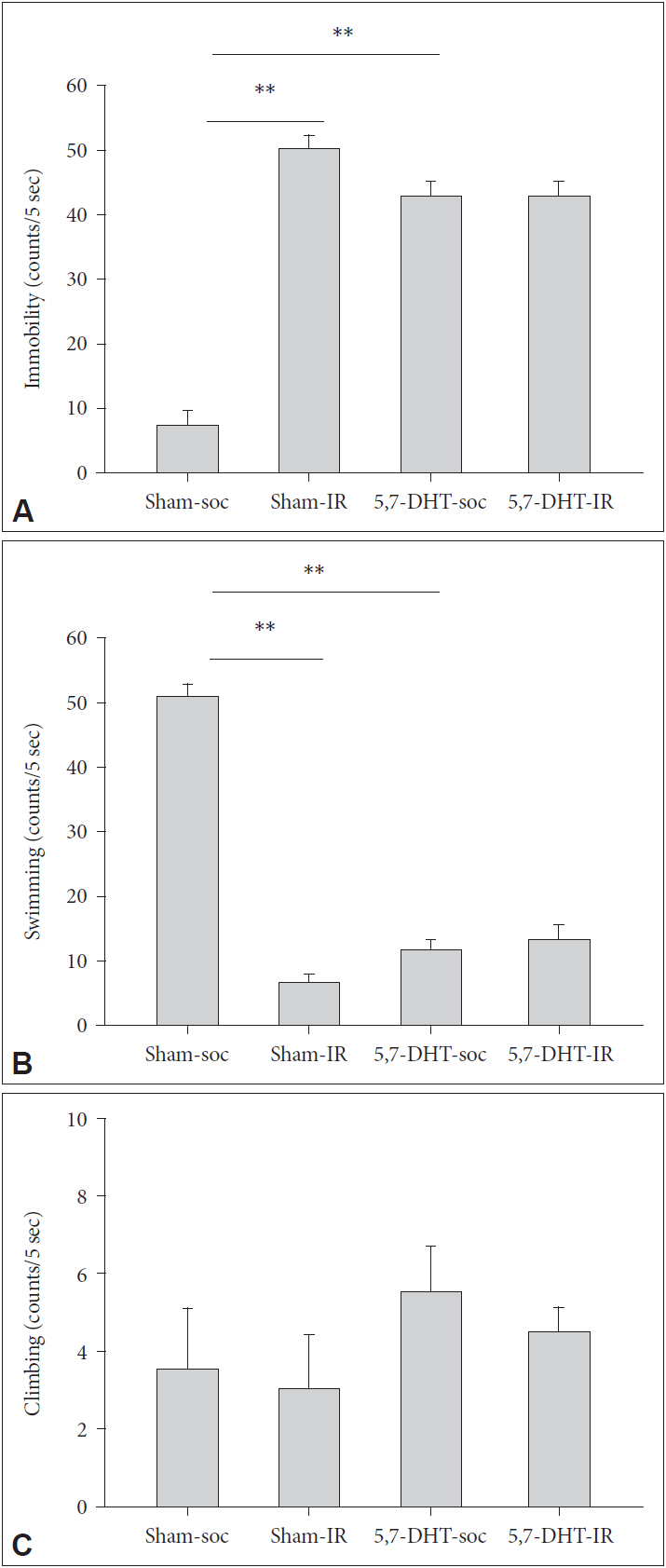
Effects of intracerebraoventricular (ICV) injection of 5-HT depletion agent, 5,7-DHT on the performance of forced swim test (FST) in rats of IR and social control (SOC) (N=5, for each group). Scored as (A) Immobility, (B) Swimming, and (C) Climbing of rats reared in social (SOC) or isolation (IR) conditions. Data are represented by mean±SEM. **p<0.01. SEM: standard error of the mean.
Effects of 8-OH-DAPT on the performance of FST
There were main effects of REARING CONDITION in which IR rats exhibited greater score of immobility [F(1,16)=9.41, p<0.01], and LESION in which 5,7-DHT caused rats more immobile [F(1,16)=8.96, p<0.01]. LESION caused rats a lower score of swimming which was more pronounced in IR rats [REARING CONDITION×LESION, F(1,16)=6.05, p<0.05] (Figure 3).
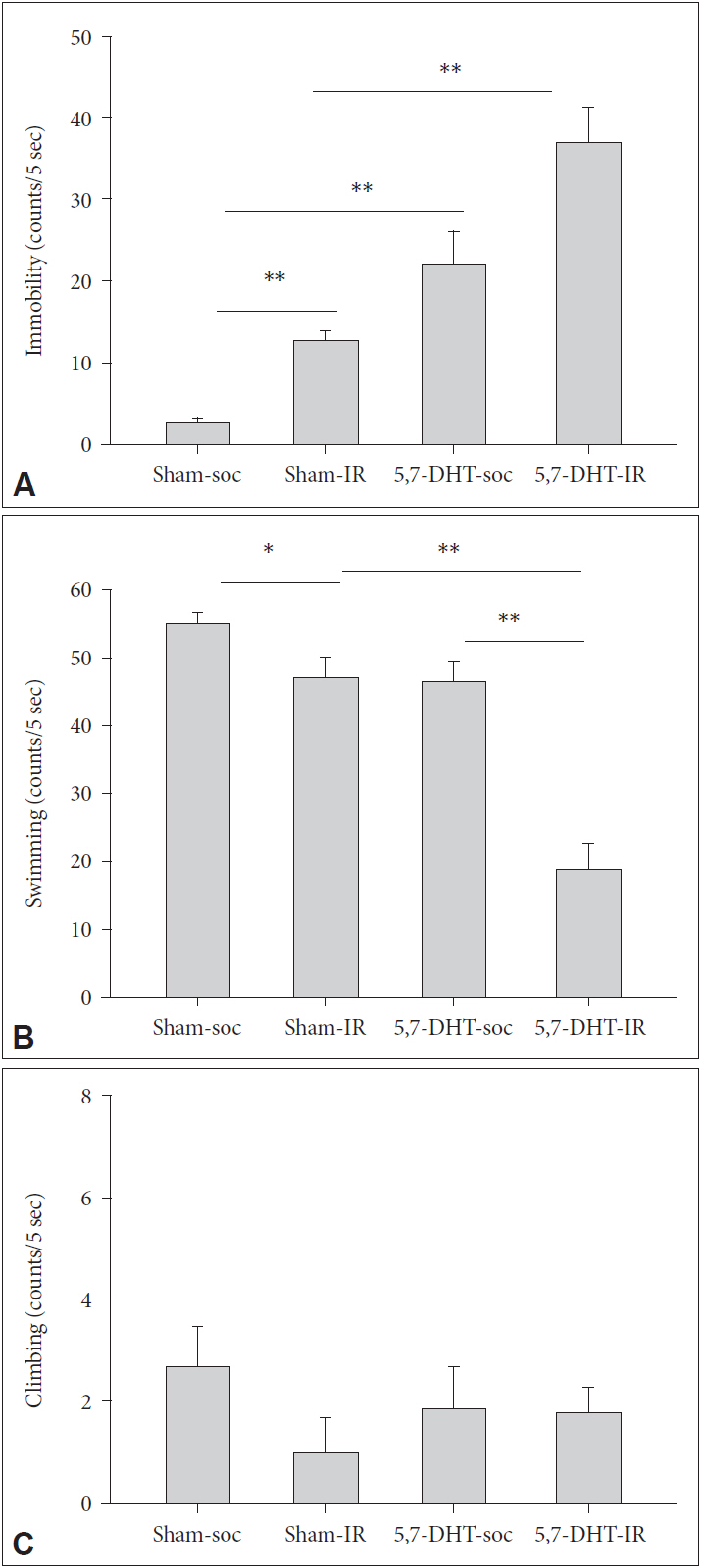
Acute effects of 8-OH-DPAT on the performance of forced swim test (FST) in rats of IR and social control (SOC) under sham and 5,7-DHT depletion conditions (N=5, for each group). Scored as (A) Immobility, (B) Swimming, and (C) Climbing of 5,7-DHT treated rats reared in social (SOC) or isolation (IR) conditions. Data are represented by mean±SEM. *p<0.05, **p<0.01. SEM: standard error of the mean.
Effects of 5,7-DHT on the expression of 5-HT1A receptors
There were no effect of REARING CONDITION×LESION on the protein expression of 5-HT1A receptors in prefrontal cortex, hippocampus, and amygdala. For prefrontal cortex, 5,7-DHT caused a main effect to increase the expression of 5-HT1A receptors [F(1,16)=4.88, p<0.05] (Figure 4).
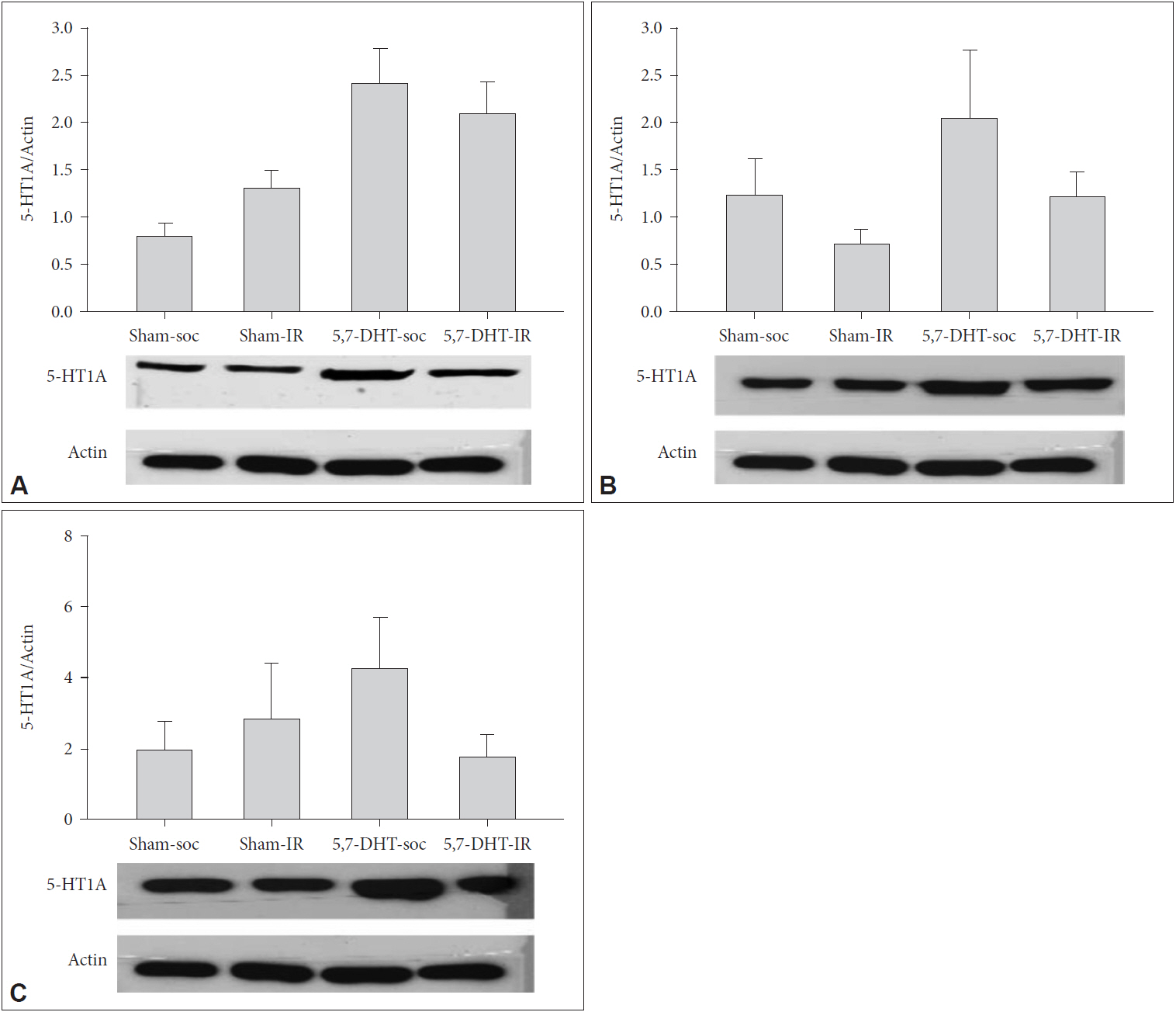
Effects of intracerebraoventricular (ICV) injection of 5-HT depletion agent, 5,7-DHT and isolation rearing (IR) on the expressions of 5-HT1A receptors of (A) prefrontal cortex (PFC), (B) hippocampus (HPX), and (C) amygdala (AMY) in rats of IR and social control (SOC) (N=5, for each group). Data are represented by mean±SEM. SEM: standard error of the mean.
Discussion
Environmental adversity during the developmental process is crucial in determining individual’s behaviors afterward [6], in which the lack of social experience in early life has been found highly relevant to animals’ depression-like profile indexed by the degree of behavioral despair as performed in FST [19]. The present study focused on the role of central 5-HT system in rats of social deprivation since weanling and found that 1) IR rats were less sensitive to the 5-HT depletion-induced immobility, 2) this effect could be reversed by acute challenge of 8-OH-DPAT, and 3) 5,7-DHT lesion increased the expression of 5-HT1A receptors in both SOC and IR rats. These results suggest the integrity of central 5-HT system is important to 5-HT1A-modulated depression profile in rats reared in social isolation. The interpretations of these findings are discussed below.
Behaviorally, in terms of developmental impact, our IR rats showed a greater immobility in FST than their SOC controls, referring a depressive sequel following long-term social deprivation [7,20]. On the other hand, 5-HT depleting lesion may cause rats to give up struggling in despairing situation of FST, this is in line with consensus that the disruption of 5-HT biosynthesis causes animals to be depressed [21]. However this phenomenon was only observed in SOC but not IR rats, the latter demonstrated a restraint ability to keep themselves not to be too immobile (Figure 2). Our data at one end supports the hypothesis that early life social deprivation may lead to depression afterward [7]. It, on the other hand, raises a possibility that individuals undergoing persistent adversity (such as a long-term lack of social attachment) might develop an ability to cope with depression [22-24].
The above finding may add to the increasing evidence that IR rats may develop a series of mediating behaviors or an adjunctive form of behavior to cope with hypothetically heightened anxiety situation, as observed in schedule-induced polydipsia [25,26] and waiting impulsivity tasks [27]. This is helpful in interpreting our findings given the fact that anxiety and depression (where the present study addressed) are very often co-existed [28] and one of the neuronal targets shared in their pathogeneses is the readjustment of the central 5-HT1A receptors, as it has been found highly implicated in both anxiety and depression [29]. In the present study, the phenomenon of less sensitivity of lesion-induced FST immobility in the IR rats became disappeared when challenged with 8-OH-DPAT, in other words, the intervention with 5-HT1A agonist seemed revive the effect of 5,7 DHT. As the lesion caused by 5,7 DHT primarily targeted at the presynaptic side, it is plausible that the effects of 5-HT1A activation involved in coping stress are associated to some postsynaptic mechanisms [30].
For the neurobiochemical aspects, 5,7-DHT in the present study increased the protein expression of 5-HT1A receptors, especially for the PFC, suggesting a upregulation of postsynaptic 5-HT1A function following the 5,7-DHT lesion although their presynaptic change seems less sensitive to be detected, as observed in the quantitative autoradiography analyses [31]. This is particularly relevant to the 5,7-DHT condition as it primarily sabotaged the presynaptic functions.
The present study has several annotations/limitations that must be addressed. First, idea of this study was to evaluate how 5-HT1A receptors react under a compromised 5-HT system. However, given the facts that 5,7-DHT intervention may also affect 5-HT2 and 5-HT3 receptors [32], it is possible that the observed 5,7-DHT effects cannot be interpreted solely by 5-HT1A receptors, particularly that 5-HT1A receptors may be also involved in the anxiety phenotype [33]. Second, lesion-induced increase of immobility observed apparently in SOC but not IR rats, it is also possible because of the ceiling effects in IR rats, i.e., they became more difficult to go higher of their immobility score. Third, in our data it seemed that the changes of 5-HT1A receptors were not directly corresponding to changes of rats’ FST score, as the behavioral output is not always explained individually by a given neurochemical index. Finally, in the present study, the 5-HT1A receptors were only acutely activated, thus the chronic effects of 5-HT1A receptors in socially isolated rats were not discussed. It is suggest to employ a chronic administration regime of 5-HT1A, for example, buspirone [34], in future studies.
In summary, our data in a way support the hypothesis that the antidepressant effect of 8-OH-DPAT is predominantly mediated by post-presynaptic 5-HT(1A) receptors [33]. We suggest that 5-HT1A may exert the key role in the IR-induced psychological outcome.
Acknowledgements
This research was supported by grants from Ministry of Science and Technology (MOST 105-2410-H-016-002-MY2), National Defense Medical Center (MAB102-83), Cheng Hsin General Hospital (CH-NDMC-106-3 and CH-NDMC-107-1), the Hualien Armed Forces General Hospital (805-C106-03,805-C107-06), and Tri-Service General Hospital (TSGHC106-107) of Taiwan.