Circadian Rhythm Hypotheses of Mixed Features, Antidepressant Treatment Resistance, and Manic Switching in Bipolar Disorder
Article information
Abstract
Numerous hypotheses have been put forth over the years to explain the development of bipolar disorder. Of these, circadian rhythm hypotheses have gained much importance of late. While the hypothalamus-pituitary-adrenal (HPA) axis hyperactivation hypothesis and the monoamine hypothesis somewhat explain the pathogenic mechanism of depression, they do not provide an explanation for the development of mania/hypomania. Interestingly, all patients with bipolar disorder display significant disruption of circadian rhythms and sleep/wake cycles throughout their mood cycles. Indeed, mice carrying the Clock gene mutation exhibit an overall behavioral profile that is similar to human mania, including hyperactivity, decreased sleep, lowered depression-like behavior, and lower anxiety. It was recently reported that monoamine signaling is in fact regulated by the circadian system. Thus, circadian rhythm instability, imposed on the dysregulation of HPA axis and monoamine system, may in turn increase individual susceptibility for switching from depression to mania/hypomania. In addition to addressing the pathophysiologic mechanism underlying the manic switch, circadian rhythm hypotheses can explain other bipolar disorder-related phenomena such as treatment resistant depression and mixed features.
INTRODUCTION
Mood disorders are a major cause of morbidity worldwide and a potentially life-threatening condition because of the risk of suicide.1,2 In response to overwhelming stressful life events, depressive mood may be considered a somewhat normal phenomenon. However, it turns problematic when mood episodes are induced by minor stressors, when episodes without apparent stressors become recurrent, and when individuals suffer a serious manic episode. Two groups of mood disorders are broadly recognized: major depressive disorder (MDD) and bipolar disorder (BD). MDD is also known as unipolar depression to distinguish it from BD. The reported prevalence rate of MDD is 5-16% and that of BD is approximately 5-6.4% in the general population.3,4 A combination of genetic, environmental, and social factors is believed to be involved in the development of mood disorders;5-7 however, the underlying pathomechanism, particularly with regard to BD, remains unclear.
The circadian system regulates daily rhythms of physiology and behavior, such as the sleep/wake cycle, body temperature, hormonal secretion, and mood.8 Interestingly, one of the most prominent clinical characteristics of BD is its rhythmicity.9 Indeed, several clinical reports have described phase advances or fast circadian rhythms in BD. For example, Kripke et al.10 reported that 5 out of 7 patients with BD exhibited fast circadian rhythms, which were slowed by the representative mood stabilizer, lithium. Furthermore, it has been reported that lithium can slow down circadian periodicity and modify circadian cycle length across species.11 In one study, compared to healthy controls, manic patients demonstrated higher nocturnal cortisol levels and an earlier timing of the nadir of the circadian rhythm.12
A considerable subset of patients with MDD or BD improve rapidly and definitely after sleep deprivation therapy.13 The social zeitgeber theory suggests that stressful life events lead to the disruption of "social zeitgebers" such as sleep schedule and mealtime, which in turn alter molecular and cellular rhythms in susceptible individuals, ultimately resulting in mood disorders.14 Social rhythm stabilization by interpersonal and social rhythm therapy is effective at reducing relapse in BD.15 Furthermore, bright light therapy is purportedly effective for treating seasonal and non-seasonal major depression as well as bipolar depression.16,17 Such clinical findings support the hypothesis that circadian rhythm dysfunction plays a causal role in mood disorders.
Recently, an excellent review by McClung18 summarized the latest data that implicate the circadian system as a vital regulator of a variety of systems believed to play a role in the development of mood disorders. Moreover, McCarthy and Welsh19 reported that circadian clock dysfunction can cause mood disorders; they suggested that non-suprachiasmatic nucleus circadian clocks such as those located in the lateral habenula, ventral tegmentum, and hippocampus could directly contribute to the pathophysiology of mood disorders.
When treating patients with BD, clinicians often encounter challenges that cannot be readily explained. For example, some patients with BD demonstrate poor response to antidepressant treatment, whereas others abruptly switch to mania or become rapid cyclers when treated with antidepressants. Such phenomena can even be observed in the same subject using the same antidepressant at different time points. A mixed state, whereby symptoms of mania and depression occur simultaneously, is another problematic occurrence. While the coexistence or rapid alternation of depressive and manic symptoms in the same episode may indicate a more severe form of BD, the definition of a mixed state is not well established and its mechanism is not well understood. We suggest that circadian rhythm hypotheses may explain the mechanism of mixed features, treatment resistant depression, and manic switching in mood disorders.
HYPOTHESES
While many hypotheses exist regarding the pathogenesis of mood disorders, we believe 3 major vulnerability factors must be considered while rating their pertinence. These include hypothalamus-pituitary-adrenal (HPA) axis hyperactivation, monoamine dysregulation, and circadian rhythm dysregulation. These 3 vulnerability factors can potentially affect each other; therefore, we put forward the triple susceptibility model of BD (Figure 1).
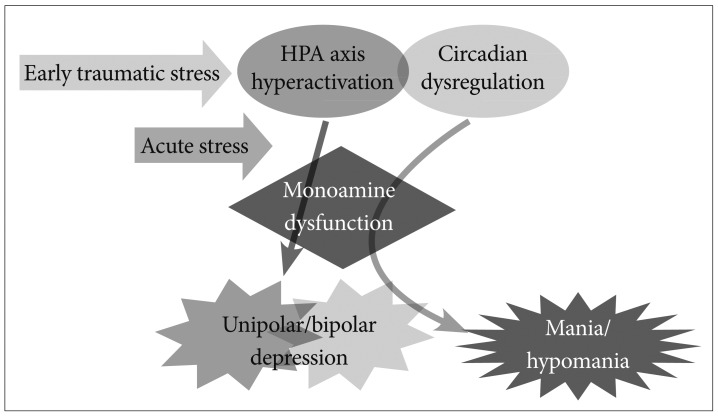
Triple susceptibility model of bipolar disorders. In addition to HPA axis hyperactivation and monoamine dysfunction, circadian rhythm dysregulation induces mania/hypomania via monoamine overactivity.
Previous studies have reported that stress during early life can cause alterations in HPA axis functioning later in life.20,21 Findings from animal studies suggest that early-life stress can produce persistent epigenetic changes that lead to hyperactivity of the HPA axis in adulthood, which in turn increases susceptibility to mood disorders.22,23 Interestingly, HPA axis hyperactivity in depression has been associated with hypercortisolemia and reduced inhibitory feedback. Studies have consistently demonstrated that glucocorticoid receptor (GR) function is impaired in MDD, resulting in reduced GR-mediated negative feedback on the HPA axis.24,25 Patients with mood disorders also demonstrate reduced GR mRNA levels in the hippocampus and frontal cortex.26,27 The crucial role of GR signaling in HPA axis regulation was confirmed in animal models of specific glucocorticoid receptor (NR3C1) knockouts.28 Compared to wild type mice, Nr3c1 gene knockdown mice exhibit increased stress-induced corticosteroid levels and impaired negative feedback with increased helplessness after stress.29 Furthermore, human genetic studies have reported that polymorphisms in GR genes are associated with mood disorders, especially BD.30-32
Monoamine dysregulation is another major hypothesis of mood disorders. Researchers have proposed that depression is caused by a functional deficiency of brain monoaminergic neurotransmitters such as norepinephrine, serotonin, and/or dopamine, whereas mania is caused by functional excess of monoamines at critical synapses in the brain.33-35 This hypothesis is based on the correlation of psychiatric symptoms and cellular actions of a variety of psychotropic agents. In the 1950s, monoamine oxidase inhibitors and tricyclic antidepressants were accidentally identified to be effective treatments for depression. Although the monoamine hypothesis received some criticism,36 it has been the major focus of pathophysiological and pharmacotherapy research for mood disorders. Individuals with an underlying vulnerability of HPA overactivity are susceptible to stress and are easily depressed following impairment of hippocampal monoaminergic neurotransmission.37,38 According to the first 2 hypotheses, individuals with a preexisting vulnerability with regard to the HPA axis and the monoamine system may be easily depressed in response to a stressful life event.
Some individuals under depression demonstrate symptoms of mania or hypomania in these situations, although this phenomenon is not clearly understood. Traditional psychoanalysts have regarded the occurrence of these symptoms as "manic defenses" wherein the patient uses it to avoid the depression after being convicted for destroying an internal object.39 We suggest that some individuals under depression switch to mania/hypomania caused by circadian rhythm instability in addition to susceptibility to HPA axis hyperactivity and monoamine system dysfunction (Figure 1).
WHAT IS CIRCADIAN SYSTEM?
Circadian rhythms are cyclic and persistent patterns of behavior, physiological changes, and mental characteristics exhibited by most living organisms on the Earth, from bacteria to humans. They occur with a 24-h period and are thus roughly reflective of the time it takes for the Earth to complete a rotation. Generally, it is believed that circadian clocks evolved in parallel with the geological history of the Earth, and that they have since been fine-tuned under selection pressures imposed by cyclic environmental changes.40 The rhythm is endogenous in humans and other organisms.41 In the absence of external time cues, it often persists for a slightly longer period than 24 h; however, in normal conditions, it is synchronized to a 24-h light/dark cycle. In humans and other mammals, light stimuli falling on the retina excite intrinsically photosensitive retinal ganglion cells, which send their axons to the suprachiasmatic nuclei (SCN). The SCN, located in the anterior hypothalamus above the optic chiasm, is considered the master pacemaker of circadian rhythms in mammals.42 Indeed, lesions of the SCN disrupt circadian oscillations lead to arrhythmicity in animals. Circadian rhythmicity can be restored by implanting fetal SCN cells into the arrhythmic host.43,44 The SCN orchestrates circadian rhythms in behavior and physiology via endocrine and neural pathways. Projections from the SCN to other hypothalamic nuclei and other brain regions control various rhythms such as sleep-wake cycles, body temperatures, and hormonal levels.45,46 Individual cells in the SCN, functioning as individual circadian oscillators, can independently regulate their own rhythm and can couple to form a master pacemaker.47 As demonstrated recently, hormonal systems that are under distinct circadian control include melatonin and the HPA axis including cortisol, the hypothalamic-pituitary-thyroid axis, and epinephrine.48,49 In addition, hormones under SCN control may serve to entrain circadian oscillations in peripheral tissues.50
WHAT HAPPENS TO THE CIRCADIAN SYSTEM IN BIPOLAR DISORDER?
Circadian system instability is observed in BD; virtually all patients possess major alterations in circadian functions such as sleep, activity, hormonal secretions, and appetite.51 Findings from various studies have revealed that patients with BD demonstrate seasonal patterns of behavior, with more manic episodes observed during the spring and summer months.52,53 This highlights a potential circadian component of the pathophysiology of BD, which is possibly influenced by alterations in the photoperiod, i.e., seasonal changes in day length. Normalization of both sleep and wake cycles and social zeitgebers are often essential for clinical recovery, whereas disruption to circadian rhythms could trigger manic episodes.54 Prevalence of mania is higher in subjects with delayed sleep phase syndrome than in controls.55 This is consistent with reports stating that compared to controls, patients with BD have a greater evening preference.56 Recently, an association between circadian gene variants and BD has been described,57-63 which may suggest that the internal regulation of circadian rhythms in BD is weak and susceptible to both internal and external influences. In contrast, genome-wide association studies (GWAS) have not identified BD-associated clock genes;64-66 however, contrasting finding was obtained upon reanalysis of 14 GWAS studies using a more focused approach. This study revealed that associations between BD-spectrum illnesses and lithium-responsiveness were enriched among core clock genes and pervasively rhythmic clock-controlled genes.67 Moreover, combining GWAS findings with the findings of convergent functional genomic studies also implicates several clock genes in BD.68
Several other experiments have indicated a link between disrupted circadian rhythms and manic behavior in mammals. For example, findings from a study that used mice carrying a mutation in the circadian gene, Clock, indicated that Clock gene mutations increase the exploratory behavior and activity of mice when exposed to a novel and stressful environment.69 Furthermore, in a forced swim test, where learned helplessness can be analogous to depression, Clock mutant mice spent significantly more time swimming in the forced swim test than wild-type mice did.69 Glycogen synthase kinase-3β has recently been implicated as a central regulator of the circadian clock,70 and this enzyme is a known target of the representative mood stabilizer lithium.71 Lithium modulates circadian rhythms by lengthening the circadian period in several organisms, including Drosophila, rodents, and humans; an effect that may be important for its therapeutic purposes.10,11,72-76 Additionally, another mood stabilizer, valproate, alters the expression of several circadian genes in the amygdala,77 whereas chronic treatment with the antidepressant, fluoxetine, increases the expression of Clock and Bmal1 in the hippocampus.78
Regarding the effects of light exposure, patients with BD appear to be more sensitive to the biological effects of light.79 The antidepressant effects of bright light therapy and sleep deprivation are more effective in these patients than in patients with unipolar depression.80,81 Bright light therapy precipitates hypomanic/manic episodes in susceptible patients,82-84 and dark therapy is effective in manic patients.85 Most successful treatments for mood disorders rely on altering the circadian cycle.86 Altogether, these findings support the view that circadian regulatory processes in the brain are important for the development and treatment of mood disorders, in particular for manic episodes.
CIRCADIAN SYSTEM AND MONOAMINE SIGNALING
Numerous studies have reported that monoamines such as serotonin, norepinephrine, and dopamine exhibit circadian rhythmicity in their levels, release, receptors, and synthesis-related enzymes.18 Recently, animal studies have suggested that there may be a link between circadian-clock components and regulation of the monoamine system.87 For example, it was observed that transcription of the monoamine oxidase A (MAOA) promoter is regulated by the clock components, Bmal1, Npas2, and Per2 in mice, and a mutation in Per2 in mice leads to reduced expression and activity of MAOA in the mesolimbic dopaminergic system. Increased levels of dopamine and altered neuronal activity in the striatum were also noted, which might have led to mania-like behavioral alterations in Per2 mutant mice.88 Spencer et al.89 reported that mPer1 and mPer2 messenger RNA levels are altered in the nucleus accumbens in response to chronic stress in mice. They also reported that selective knockdown of both mPer1 and mPer2 via RNA interference in this region induces anxiety-like behavior.
Dominant-negative mutant (ClockΔ19) mice have a behavioral profile that is very similar to manic symptoms such as hyperactivity, low anxiety/depression behavior, and increased reward seeking.90,91 Interestingly, ClockΔ19 mice have increased dopamine synthesis and activity as well as increased tyrosine hydroxylase expression in the ventral tegmental area (VTA).90,92 Furthermore, it was observed that the majority of mania-like behavior in these mice was reversed with chronic lithium treatment.91 Lithium also restored normal levels of VTA dopaminergic activity in the Clock mutants.92
The circadian system regulates monoaminergic activity in various brain regions that control mood and behavior through local expression of clock genes as well as indirect connections originating in the SCN. The SCN projects monosynaptically to multiple hypothalamic nuclei, which subsequently communicate with regions that synthesize dopamine, serotonin, and norepinephrine.18
ANTIDEPRESSANT-RESISTANT DEPRESSION AND MANIC SWITCHING
A considerable number of patients diagnosed with unipolar depression are later diagnosed with BD. A survey reported that 48% of patients with BD consult 3 physicians before they are correctly diagnosed, while 38% of the patients take more than 10 years for a correct BD diagnosis.93 Early and precise diagnosis of BD and early effective treatment intervention are pivotal considering the evidence that antidepressants sometimes, if not always, stimulate poor treatment response and manic switching or rapid cycling in patients with BD in depressive episodes.94
Previous studies have suggested that monoaminergic antidepressants might increase the risk of manic switching95,96 or rapid cycling.97-99 A strong association between poor antidepressant response and a subsequent change in diagnosis to BD has also been reported.100 In a controlled trial, antidepressant-induced manic switching was observed in up to 50% of subjects not treated with mood-stabilizing medication.101 However, despite these findings, the mechanism underlying manic switching and antidepressant resistance remains elusive. Interestingly, decreased amount of total sleep and shortened latency of rapid eye movement sleep occur at the time of the switch to mania,102,103 and disturbed sleep patterns are also associated with circadian fluctuations in body temperature and hormone secretion as well as catecholamine increase.9 Some treatment methods for depression have a phase-advancing effect on circadian rhythms. Morning light therapy leads to a phase advance in biological rhythms in seasonal affective disorder, and phase alignment either through light therapy or melatonin treatment is necessary for recovery.104 Antidepressants such as fluoxetine produce a phase advance in the firing of SCN neurons in rats.105 These findings implicate changes in sleep patterns and other circadian disturbances in the switch process.
We hypothesize that manic switching and treatment resistance to antidepressants in BD can be explained by circadian rhythm hypotheses. A percentage of patients with recurrent depression may also possess a covert form of BD. Switching may relate to the circadian rhythmic phase, wherein individuals experience depression upon phase delay and slowing-down of the circadian rhythm, whereas they come out of the depressive state upon quickening up of the circadian rhythm and phase advancement. Even though patients with depression act as if they are in the same depressive state, they may have different varieties of circadian phase disorders. This is depicted in Figure 2. Furthermore, when the patient's mood is on the downhill slope leading to depression during the slower circadian phase, they do not respond to antidepressant treatment; however, when the mood is on the uphill slope towards euthymic or elated mood and the circadian rhythm is faster, patients are more responsive and manic switching can be rendered.
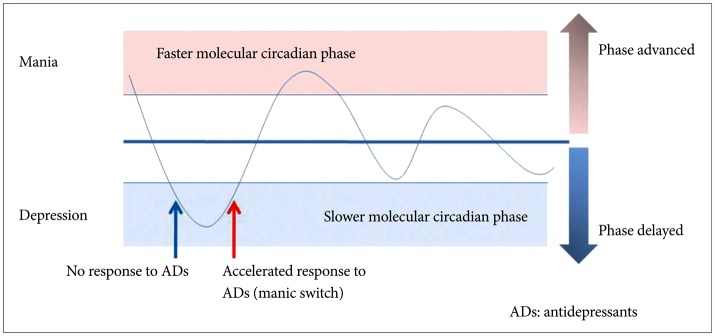
When the mood state is on the downhill slope in the slower circadian phase leading to depression, antidepressants is not effective. In contrast, when the mood on the uphill slope is faster circadian rhythm leading to euthymia or mania, antidepressants can induce very fast response or sometime render manic switch.
HOW CAN MIXED STATES BE EXPLAINED?
Mixed state, also known as dysphoric mania, agitated depression, mixed features, or a mixed episode, is a condition during which features of mania and depression occur simultaneously. However, the definition and pathophysiology of mixed states are not well established. Occasionally, the mood alternation can take place over the course of a day or more, but in other instances, alternations between depressed and manic manifestations may occur over a few hours or even switch back and forth within a few minutes.
Kraepelin106 suggested that some mixed states are transitions between depressive and manic episodes. Transitions between manic or depressive states are common, as previously demonstrated in a longitudinal study.107 Polyphasic episodes, in which state changes occur without a euthymic mood period, resemble mixed states.108 Interestingly, Sit et al.17 reported that mixed states were induced by morning light treatment in women with bipolar depression; however, midday light therapy did not induce mixed states. This finding suggests that abrupt phase advancement by morning light therapy underlies the pathomechanism of mixed states.
Recently, in an animal study, Mukherjee et al.109 reported that Clock knockdown, specifically in the VTA, results in hyperactivity and a reduction in anxiety-related behavior, which is phenotypically similar to Clock mutant mice. However, in their study, VTA specific knockdown also resulted in a substantial increase in depression-like behavior, creating an overall mixed state. They further reported that Clock knockdown in the VTA also altered the circadian period and amplitude, suggesting a role for VTA Clock gene in the regulation of circadian rhythms.
We suggest that circadian rhythm hypotheses may explain the pathophysiology of mixed states. Therefore, for example, if a patient with BD has a serious course of illness and if his mood shifting occurs without a substantial period of mood stability, the transitional period between slow and fast circadian phases may result in unstable mood leading to a mixed state of depressive and manic symptom at the same time (Figure 3). The rapid transitional period of biological rhythm does not persist for long time because of the existing sustained endogenous circadian rhythm. For example, we experience jet lag because of alterations to the body's circadian rhythms resulting from rapid long-distance airplane travel.
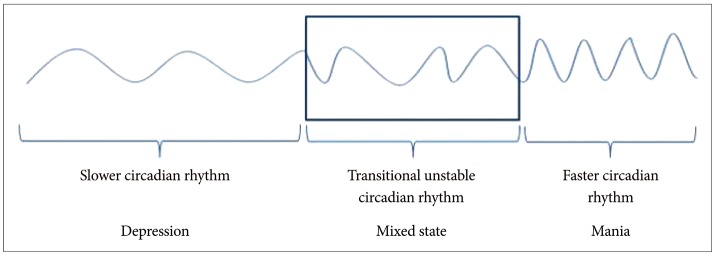
In case of mood shift happening without a substantial period of stable mood, i.e., during the transitional period between slower circadian rhythm of depression and faster one of mania, mood can be unstable and a combination of depressive and manic symptom at a same time.
CONCLUSIONS
Circadian rhythm hypotheses are regarded as a promising explanation for BD development. Circadian rhythm instability, imposed on the dysfunction of HPA axis and monoamine system, may in turn increase individual susceptibility for switching from depression to mania/hypomania. Furthermore, different circadian phases may explain altering treatment response to antidepressants as well as the phenomena of mixed features. Recent data implicates the circadian system in the development of mood disorders. We suggest that dysregulation of the circadian system regulates the mechanisms of mixed features, antidepressant treatment resistance, and switching in mood disorders.
Acknowledgments
We wish to thank Dr. Daniel F. Kripke for his comments on the manuscript. This study was supported by the Korea Health 21 R&D Project funded by the Ministry of Health & Welfare, Republic of Korea (HI11C1901).