Promising Neuroimaging Biomarkers in Depression
Article information

Abstract
The neuroimaging has been applied in the study of pathophysiology in major depressive disorder (MDD). In this review article, several kinds of methodologies of neuroimaging would be discussed to summarize the promising biomarkers in MDD. For the magnetic resonance imaging (MRI) and magnetoencephalography field, the literature review showed the potentially promising roles of frontal lobes, such as anterior cingulate cortex (ACC), dorsolateral prefrontal cortex (DLPFC) and orbitofrontal cortex (OFC). In addition, the limbic regions, such as hippocampus and amygdala, might be the potentially promising biomarkers for MDD. The structures and functions of ACC, DLPFC, OFC, amygdala and hippocampus might be confirmed as the biomarkers for the prediction of antidepressant treatment responses and for the pathophysiology of MDD. The functions of cognitive control and emotion regulation of these regions might be crucial for the establishment of biomarkers. The near-infrared spectroscopy studies demonstrated that blood flow in the frontal lobe, such as the DLPFC and OFC, might be the biomarkers for the field of near-infrared spectroscopy. The electroencephalography also supported the promising role of frontal regions, such as the ACC, DLPFC and OFC in the biomarker exploration, especially for the sleep electroencephalogram to detect biomarkers in MDD. The positron emission tomography (PET) and single-photon emission computed tomography (SPECT) in MDD demonstrated the promising biomarkers for the frontal and limbic regions, such as ACC, DLPFC and amygdala. However, additional findings in brainstem and midbrain were also found in PET and SPECT. The promising neuroimaging biomarkers of MDD seemed focused in the fronto-limbic regions.
INTRODUCTION
Major depressive disorder (MDD) is an important psychiatric illness with substantial impacts in many perspectives of function and life quality. The functional impairments were correlated with the clinical symptoms [1]. The physical, psychological and social domains of life quality is inter-related to functional status [2]. MDD symptoms would interfere with concentration, motivation and cognitive functions, which would cause patients to have daily functional impairments [3]. Therefore, it is important for scientists and clinicians to understand the pathophysiology of MDD. Many articles about the structural and functional pathophysiology of MDD mentioned the possible roles of fronto-limbic network [4-7]. The fronto-limbic network seemed to originated from the initial “limbic-corticalstriatal-pallidal-thalamic tract” model for MDD [8]. Recently an evolved MDD model mentioned the importance of ventromedial prefrontal cortex, anterior cingulate cortex (ACC) and lateral parietal cortex, which might be associated with self-referential problems and negative ruminations [9].
In the pathophysiology model, ACC seemed to play a significant role in MDD. ACC was also responsible for important functions of MDD, such as the attention, problem solving, motivation, decision making [10-12]. Two components of ACC, affective and cognitive subdivisions [11,13], might be associated with functional impairments in MDD. The affective subdivision was connected with limbic regions, such as the amygdala and brainstem, to modulate the emotions [14]. The cognitive subdivision was involved in cognitive processing of MDD [13]. The cognitive and affective components of ACC might have important roles in the pathophysiology of MDD [15,16]. Therefore the MDD pathophysiology biomarkers might include the ACC. In addition to the ACC, other regions of frontal lobe, such as dorsolateral prefrontal cortex (DLPFC) [17-20], would be crucial for the cognitive control of depression symptoms. The emotional responses and depression symptoms might be originated from the limbic regions, such as the hippocampus and amygdala [8,18,21-25]. From the viewpoint of cognitive control of emotional responses, the fronto-limbic network or circuit would be the state-of-art model to explain the influences of cognition impairments and the exaggerated negative emotion responses from the limbic regions in MDD.
However, the promising neuroimaging biomarkers in MDD were still not conclusive according to the extent and focus of current study. In this article, I would focus on the literature review of neuroimaging in the study of pathophysiology of MDD, including the magnetic resonance imaging (MRI), magnetoencephalography (MEG), near-infrared spectroscopy (NIRS), electroencephalography (EEG), positron emission tomography (PET), single-photon emission computed tomography (SPECT). The potentially promising biomarkers for the neuroimaging perspectives would be discussed and summarized to establish a possibly conclusive biomarker model for MDD.
MDD biomarkers revealed by MRI and MRS
The MRI part of neuroimaging biomarkers would include the magnetic resonance spectroscopy (MRS), structural and functional MRI. The MRS can measure the neural metabolites inside the brain, such as the N-acetyl aspartate and choline. The MRS study of adolescent melancholic MDD depression patients had significant correlations between tryptophan metabolites and neurotoxic metabolites in the limbic regions, such as the striatal areas [26]. A study using functional MRI integrated classifiers showed better predictability and accuracy for the detection of biomarker areas involving the emotion processing area [27]. The visual processing of emotional faces in MDD showed limbic activations, especially in the hippocampus [28]. The pre-school children MDD also had heightened responses to sad faces in the limbic regions, such as the amygdala [29]. The disrupted amygdala activity during face emotion identification in depressed children around 4–6 years old, which suggested the potential biomarker role of amygdala in endogenous depression [30]. The genome-wide neuroimaging study showed a link between a potential candidate of specific genotype and hippocampus volume, which suggested the complex relationship between genotype and MRI biomarkers in a more comprehensive model for biomarkers in MDD [31]. The baseline brain activities would be fluctuated in the hippocampus of MDD patients with suicide ideation [32]. The hippocampus was also the state-dependent biomarker region for antidepressant treatment in MDD [33]. The recurrent MDD patients also had lower gray matter volume (GMV) in the left hippocampus [34]. The hippocampus-related neural network, including the DLPFC, was also altered in the young subjects with family history of MDD, especially during a memory task [35]. The hippocampus tail volumes were relatively larger in MDD and the hippocampus tail volumes would be associated with clinical remission after antidepressant treatment [36].
The frontal dysfunction has also been replicated in many MRI studies of MDD. A MRS study showed an increased choline/creatine ratio in the ACC of MDD patients, which might be a possible diagnostic aid tool [37]. A review article of MRS study in MDD showed higher choline levels in the frontal lobe of depression patients [38]. MDD could also be differentiated from the panic disorder using regional homogeneity pattern in the left ACC [39]. The meta-analysis of GMV studies in MDD also revealed the GMV alterations in bilateral ACC [40]. A meta-analysis of resting-state functional MRI studies suggested a possible pattern of diagnosis classification according to the hyperactivity/hyper-connectivity between ACC and other prefrontal regions for the external-directed cognition [41]. The alterations in cortisol awakening responses, which would influence the hypothalamic-pituitary-adrenal axis homeostasis in MDD, were also associated with reduced GMV and increased stress-related brain activities in the ACC [42], which also corresponded to another GMV study in MDD with panic disorder [6]. A machine-learning algorithm study showed the crucial roles of areas for the guilt-selective neural signature, such as the ACC, hippocampus and thalamus [43]. The regional homogeneity of ACC might be associated with illness duration and depression severity in MDD patients [44]. The higher fluctuations of resting-state brain activity in the subgenual ACC of MDD patients would be significantly reduced after electroconvulsive therapy. In addition, the subgenual ACC activity would predict the treatment responses with the reductions in the connectivity between ACC, hippocampus and other prefrontal regions [45]. In addition, the variants of ACC thickness would predict the responses to electroconvulsive therapy [46], which corresponded to that dorsal part of ACC (cognitive subdivision of ACC) volume could predict the long-term clinical outcome trajectory [47] and prediction of rostral ACC thickness for the treatment response to transcranial magnetic stimulation in MDD [48]. The micro-integrity of cingulate bundle also predicted the treatment responses of MDD patients for antidepressants [49]. The blunted response of ACC during consummation phase and that of hippocampus during effort phase of functional MRI reward task also predicted the high risk for adolescent MDD [50]. A review article suggested that the pharmaceutical interventions would relieve functional connectivity alterations in the fronto-limbic, default mode network and parieto-temporal regions. The psychotherapeutic interventions have also been showed to influence the functional connectivity within the fronto-limbic network. The electroconvulsive and magnetic stimulation therapy would also alter the ACC, DLPFC and default mode network [51]. The depression severity was associated with elevated connectivity between fronto-limbic regions, such as ACC, amygdala and hippocampus, in a resting-state functional MRI study [52]. The distinct differences of functional connectivity in the fronto-limbic network between suicide ideators and suicide attempters were also found in the resting-state functional MRI study [53]. A meta-analysis of machine learning studies in MDD also confirmed the importance of ACC for the treatment prediction and the crucial role of fronto-limbic network for the pathophysiology of MDD [54]. A review of structural MRI studies in MDD also supported the important roles of fronto-limbic network for the pathophysiology [55]. The white matter tract study in MDD also showed the alterations in the micro-integrity of superior longitudinal fasciculus, which connected the frontal regions with the limbic regions and temporal regions [56,57]. It also supported the fronto-limbic model for the biomarker regions in MDD. The core region of the fronto-limbic network might be the ACC [58].
The magnetic seizure therapy-related cortical inhibitions of frontal cortex were the significant indicators of remission for suicidal ideation [59]. The prefrontal circuit dysfunction, such as the hypo-activations of DLPFC during the working memory updating and conscious negative emotion processing tasks of functional MRI experiments [60]. The prefrontal cortex-related cognitive control and emotion regulation abilities would be associated with resilience factors in the treatment response [61]. The persistent alterations of anterior subnetwork of default mode network, which also included frontal regions, were observed in the MDD patients after antidepressant treatment [62]. The implantation of deep brain stimulation target and the transcranial repetitive magnetic stimulation in the left DLPFC would relieve the depression symptoms in the treatment-resistant MDD adolescents [63]. The DLPFC was also a popular treatment target of brain stimulation in MDD [64]. The DLPFC and dorsal ACC cognitive subdivision could form a cognitive control network. The attenuation of intrinsic activity in cognitive control network would predict the treatment response in remitted MDD patients [65]. The transcranial magnetic stimulation in the DLPFC would also trigger higher activities of subgenual ACC in the treatment of MDD patients, which would modulate the cognitive control network [66]. The functional MRI-related global brain signal regression was more significant in the DLPFC in treatment-resistant MDD patients, which could be normalized by ketamine infusion treatment [67]. The non-remitted late-life MDD patients had significantly greater reductions in the GMV of orbitofrontal cortex (OFC) than remitted late-life MDD patients and controls [68]. The GMV of OFC also predicted the antidepressant treatment responses in MDD [34]. The deficient inhibition of return for emotional materials was also altered in the OFC and hippocampus-related activity and functional connectivity of MDD patients, which suggested that OFC might be specific for remitted responses and hippocampus might be specific for depressive phase [69].
However, some studies were still against the significant roles of fronto-limbic regions as biomarkers of MDD. A MRS study showed that N-acetyl aspartate levels showed no significant changes in the DLPFC and amygdala of first-episode MDD patients after antidepressant treatment [70]. A meta-analysis showed a lack of increases in the brain-derived neurotrophic factor after non-invasive brain stimulation in the DLPFC of MDD patients [71]. In addition, the hippocampus enlargement might not represent a sole predictor for the treatment response to electroconvulsive therapy [72].
In summary, for the MRI and MRS study in MDD, the most promising biomarkers might be the ACC. The other prefrontal regions, such as DLPFC and OFC, may cooperate with ACC to contribute to the inhibition of emotional response and emotional memory via the cognitive control network. The emotional response and memory might be originated from the limbic regions, such as the amygdala and hippocampus. The model of neuroimaging biomarkers in MRI and MRS may still exist in the architecture of fronto-limbic model. In addition, the prediction of treatment response seemed to be associated with fronto-limbic regions, especially for the ACC and hippocampus. However, there are still many efforts needed to clarify the highly promising biomarkers of MRI and MRS study in MDD, especially using the longitudinal study with randomized controlled design to investigate the specific biomarker regions for different kinds of therapeutic interventions, such as the pharmacological treatment, psychotherapeutic intervention and brain stimulation therapy.
MDD biomarkers revealed by MEG
The MEG can measure the brain activities using the algorithm to calculate the magnetic field caused by the electricity of brain activities to obtain the neuroimaging. The amount of MEG studies in MDD biomarker searching is relatively fewer than that of MRI studies. The electroconvulsive therapy might activate the parieto-temporal regions in the MEG of responding MDD patients, which suggested that the parietal lobe would influence the clinical response via the information processing and attention modulation [73]. Another MEG study in drug-free MDD patients with high suicide risk showed the impaired phase-amplitude coupling in the limbic regions, such as the caudate and thalamus. The impaired coupling might influence the DLPFC to cause the alterations in the executive function and working memory [74]. The higher scores of Lempel-Ziv complexity on MEG have been reported in MDD patients when compared to controls [75]. A multimodal imaging of MDD patients, including MEG, showed that the subgenual ACC and amygdala might be the biomarker of MDD pathophysiology and ketamine treatment in MDD [76]. Another study showed that alpha connectivity between subgenual ACC and DLPFC would predict the better response of antidepressant treatment [77]. The resting-state MEG study showed that hyper-interwines and hyper-integrates within the DLPFC and ACC might represent the MEG endophenotype of MDD [78]. The ketamine treatment in MDD also would modulate the subgenual ACC-related connectivity network, including the limbic regions [79]. The MEG-individual component analysis study also supported that subgenual ACC would be the core of biomarker network, which included the hippocampus and amygdala [80]. The MEG dynamic causal modeling study demonstrated that DLPFC might be incapable to inhibit the amygdala and the subsequent enhanced amygdala-ACC and ACC-DLPFC bottom-up signals would be associated with MDD symptoms [81]. In spite of possible inconsistences, the major architecture of fronto-limbic biomarker network may be replicated in MEG study, especially the crucial role of ACC has been proved again.
MDD biomarkers revealed by NIRS
The NIRS use the near-infrared region of spectrum to measure the blood oxygenation, blood flow and brain activities. The MDD with suicidal ideation showed smaller hemodynamic changes in the DLPFC and OFC during verbal fluency test in the NIRS study. In addition, the hemodynamic changes in the DLPFC and OFC were negatively correlated with the severity of suicidal ideation. The DLPFC and OFC might represent the neural substrate for suicidal ideation [82]. The blood flow in the frontal lobe would also be negatively associated the depression severity in another NIRS study [83]. Another study of NIRS in the dexamethasone suppression test of MDD patients showed that value of the center of gravity at the frontal lobe would be significantly greater in the non-suppressor group, which suggested that frontal lobe NIRS values might be representative of MDD patients with cortisol suppression impairments [84]. The emotional face recognition task in MDD also revealed the importance of left prefrontal cortex due to the alterations of median value and ratios of oxygenated hemoglobin/deoxygenated hemoglobin, which suggested the weaker hemodynamic oscillations in the prefrontal cortex of MDD patients [85]. The emotional Stroop task in MDD also found that an evoked wave in the left upper frontal cortex might be inversely correlated with the severity of depression [86]. The antidepressant treatment seemed to influence the left inferior frontal cortex and temporal gyri in MDD patients, which suggested that prefrontal hemodynamic alterations might be the predictor of clinical responses to antidepressant treatment [87]. A 1.5-years follow up study also confirmed the important role of frontal lobe in the state-dependent and trait biomarkers, which might be the inferior frontal cortex and middle frontal cortex respectively [88]. Several studies of NIRS also supported the crucial role of frontal lobe or prefrontal cortex in the pathophysiology of MDD and might represent the biomarker for depression [89-92]. It seemed that most NIRS studies in MDD emphasized the role of potentially promising biomarker in the frontal regions. The fronto-limbic network in the NIRS field of MDD probably would be more focused on the frontal component within this network.
MDD biomarkers revealed by EEG
The EEG is a traditional tool to understand the electric activities accompanied with brain activities. The cortical inhibition EEG signals, such as the N100 and LICI, in the DLPFC and other frontal regions might predict the treatment response to the magnetic seizure therapy. It suggested that stronger baseline inhibitory neurotransmission may represent the indicators [59]. The EEG can assess the arousal level due to dysregulation of inner tension and might assist to decide which kind of patients might be responding to psychostimulant treatment [93]. In addition, the vigilance regulation pattern of EEG has been applied to identify the MDD patients [94]. The MDD responders of repetitive transcranial magnetic stimulation showed higher baseline and post-treatment working memory-related fronto-midline theta power and connectivity than non-responders [95]. The alpha, theta and gamma oscillation waves of EEG can provide information on depressive states and recovery. The gamma rhythms can distinguish MDD patients from controls and bipolar disorder patients. In addition, the gamma rhythms can predict the treatment response of pharmacological and non-pharmacological treatment [96]. Reduced gamma rhythms in the rostral ACC has been suggested as the biomarker for MDD [97]. The resting-state EEG gamma power and theta-gamma coupling in frontal region might predict the treatment response of repetitive transcranial magnetic resonance treatment and the accompanied increases of gamma power in the frontal region would occur after treatment [98]. The antidepressant responders of MDD had greater theta current density in the rostral ACC and OFC than non-responders [99]. The EEG alpha asymmetry with greater right-lateralized activities has also been proposed as a kind of biomarker to identify MDD [100]. The metaanalysis of EEG sleep pattern in MDD showed that increased rapid eye movement density and shortening slow wave sleep might represent the EEG sleep biomarkers of MDD [101]. The REM sleep prefrontal theta cordance of MDD patients was also positively associated with the improvements in depression symptoms. In addition, the greater prefrontal theta cordance of REM sleep EEG might represent the biomarker for antidepressant treatment in MDD patients [102]. Depressed adolescents also exhibited significantly increased slow wave activities of frontal regions during sleep, which were positively correlated with morbid thoughts. It represented the cortical regions of intense use or restructuring might be influenced in MDD [103]. The prefrontal theta cordance, alpha and delta hemispheric asymmetries might also predict the antidepressant treatment in MDD [104]. The theta cordance in the DLPFC and central brain regions also could predict the treatment response of brain stimulation [105].
In summary, due to the technique limitations of EEG, the findings would be localized on the cortex surface. The prefrontal cortex, such as ACC, DLPFC and OFC, might be crucial for the biomarker detection of MDD. The EEG biomarker characteristics of the prefrontal cortex included the theta density, theta cordance, alpha rhythm asymmetry, delta rhythm asymmetry, gamma rhythm and sleep EEG. The frontal part of fronto-limbic network has been proved in the EEG findings. However, the diversity of EEG parameters might be needed to be unified and processed to develop an applicable EEG model to identify MDD patients and the biomarker of treatment response in MDD.
MDD biomarkers revealed by PET and SPECT
The PET and SPECT can detect the neurotransmitter level using the radioligand labeling the brain region with targeted neurotransmitter receptors. The neuroticism personality, which were highly associated with MDD, was positively correlated with fronto-limbic serotonin 5HT-2A receptor binding potential. The fronto-limbic serotoninergic dysregulation might be vulnerable to MDD [106]. The glucose metabolism study showed different modulation of brain glucose metabolisms by cognitive behavioral therapy and antidepressant [107]. A serotonin PET study showed that the serotonin transporter binding potential was significantly lower in the forebrain, brainstem and putamen of MDD patients with suicide attempts. It also supported the serotoninergic dysregulation in MDD patients and suicide [108]. Male MDD patients had significantly higher serotonin 5HT-1A binding potential over the raphe nuclei in the brainstem, DLPFC, ACC, other frontal regions, amygdala and hippocampus [109]. The cerebral amyloid PET study showed that cerebral amyloidosis would occur in the hippocampus of MDD patients with mild cognitive impairment [110]. The ketamine therapy in MDD would be associated with subgenual ACC glucose metabolism changes, which might suggest the crucial role of ACC for the glutamate modulation in the treatment of MDD [79]. The decreased binding of serotonin transporter in the amygdala has also proposed as the potential biomarker for remission after antidepressant treatment [111]. The binding potential of the neuroinflammation protein in subgenual prefrontal cortex and ACC might be increased in the unmedicated MDD patients, which also supported the crucial role of ACC from the viewpoint of neuro-inflmmation hypothesis in MDD [112]. The subcallosal ACC glucose metabolism was significantly higher in non-responders than remitters after cognitive behavioral therapy and antidepressant treatment [113]. The light therapy can decrease the serotonin transporter binding potential in the ACC of seasonal MDD during the winter season, which also supported the importance of ACC in the seasonal impacts of MDD via the serotoninergic regulation [114]. The density of monoamine oxidase A, a kind of enzyme to metabolize the serotonin, was found to be greater in the prefrontal cortex and ACC of patients with postpartum MDD [115]. It emphasized the crucial role of ACC and frontal regions in the pathophysiology of MDD, including the seasonal and postpartum subtypes. The deep brain stimulation also reached remission in the treatment-resistant MDD patients with higher baseline prefrontal regional cerebral blood flow, which also suggested the predictive biomarker of prefrontal cortex for deep brain stimulation [116].
The SPECT guided transcranial magnetic stimulation in the left DLPFC also improved the MDD symptoms, which also supported the DLPFC as the treatment biomarker for MDD [117]. The lower serotonin transporter binding potential in the midbrain using SPECT modality would also be associated with heritability of MDD [118]. The attenuation of negative attention bias in MDD after antidepressant treatment might be associated with serotonin transporter occupancy in the amygdala of a SPECT study in MDD, which suggested that antidepressant can relieve limbic elevated activities [119]. The distribution volume ratio of serotonin transporter would be lower in the basal ganglia and midbrain [120]. Another study of cerebral blood flow using SPECT also observed the decreased blood flow in the prefrontal cortex, especially the ACC, DLPFC and OFC, which also supported the critical role of prefrontal cortex in the blood flow biomarker of MDD [121]. Other studies of SPECT in MDD seemed to focus on the midbrain and found the alterations in serotonin transporter occupancy of midbrain [122-124].
In summary, the PET studies seemed to emphasize the ACC and other prefrontal cortex, such as the DLPFC and OFC in the biomarker search of MDD, especially for validation of the neuroinflammation theory, deep brain stimulation, antidepressant treatment effects, psychotherapy effects, seasonal effects and postpartum effects. The findings of limbic regions, such as amygdala and hippocampus, combined with the findings in the brainstem and midbrain, could be validated for the biomarkers of cognition impairment, antidepressant treatment and psychotherapy effects. For the SPECT study, most findings seemed to be focused on the midbrain, especially for the serotoninergic biomarker search in MDD. The frontal regions of fronto-limbic network might play a role in the biomarker of decreased cerebral blood flows. However, most studies of PET and SPECT were based on the region-of-interest method, which might be associated with selection bias and would influence the interpretations of the study results.
CONCLUSION
From the above literature review, the fronto-limbic network seemed the promising neuroimaging biomarkers in MDD, especially for MRI, MRS, MEG and PET study (Figure 1). The most promising region may be the ACC. The importance of frontal regions has also been confirmed in NIRS and EEG study. The frontal areas might inhibit the emotional responses from the limbic regions, such as amygdala and hippocampus. Further multimodal imaging study would be needed to elucidate the neuroimaging biomarker issue in MDD.
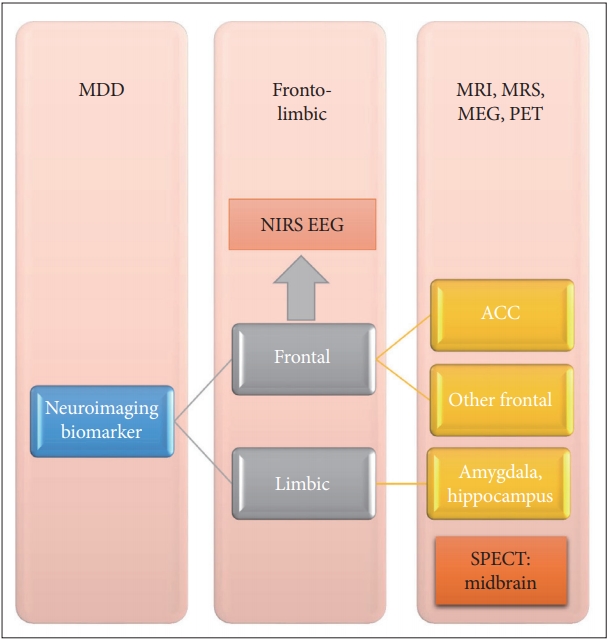
The fronto-limbic network and promising neuroimaging biomarker in MDD. The MRI, MRS, MEG, and PET study mentioned all the important parts involving the frontal and limbic regions. The most promising biomarker would be ACC. The NIRS and EEG also supported the crucial role of frontal regions. SPECT study revealed the clues of midbrain biomarker, which might be associated with the region-of-interest method. ACC: anterior cingulate cortex, EEG: electroencephalography, MEG: magnetoencephalography, MRI: magnetic resonance imaging, NIRS: near-infrared spectroscopy, PET: positron emission tomography, SPECT: single-photon emission computed tomography.
Notes
The author has no potential conflicts of interest to disclose.